|
|
|
|
| Livestock Research for Rural Development 20 (9) 2008 | Guide for preparation of papers | LRRD News | Citation of this paper |
Twenty-five (25) pregnant adolescent Djallonké (West African Dwarf) ewes were offered a basal diet of Gliricidia sepium and Brachiaria decumbens and supplemented with a single concentrate whose intake was restricted in early (T1), mid (T2) or late (T3) pregnancy, unrestricted throughout (T4) or restricted throughout (T5) pregnancy. Dry matter intake was generally highest in mid, lower in late and lowest in early pregnancy (P<0.05). Ewes in T4 which were supplemented throughout gestation gained the highest weight during pregnancy (P<0.05) but did not give birth to heavier lambs than those which were not supplemented in one stage of pregnancy (P>0.05). High weight gains during pregnancy were associated with a paradoxical phenomenon of impeded placental growth and function which affected lamb birth weight. The relationship between placental weight and lamb birth weight was significantly positive (r = 0.6489; P<0.05). Because dry matter intake is generally highest in mid pregnancy (P<0.05), rapid maternal growth can be checked with imposition of dietary restrictions in that stage of pregnancy to enhance placental growth and subsequently lead to improved lamb birth weight in adolescent ewes. Dietary restriction in only one stage of pregnancy did not generally affect total plasma protein (Tp) concentration (P>0.05) but the latter was significantly correlated with lamb birth weight (r = 0.93; P<0.05). The study explains why adolescent ewes may grow very well during pregnancy and yet give birth to lambs with poor birth weights.
Keywords: dietary restriction, late pregnancy, mid pregnancy, nutrition
In sheep, the efficiency of production depends largely on the reproductive performance of the ewe while the most critical phase of the reproductive cycle that is most susceptible to nutritional alterations is gestation (Webb et al 1999). The interaction between nutrition and reproduction therefore constitutes an important interface of livestock production that determines the success or failure of any livestock enterprise. Though the nutrition-reproduction interaction is complex and responses are often variable and inconsistent (O’Callaghan and Bolland 1999), inappropriate maternal nutrition has been reported as one of the major extrinsic factors programming lamb birth weight (Robinson et al 1994). The significant role of maternal glucose in ovine utero-placental nutrition has already been extensively investigated (Wallace et al 1997; Robinson et al 1999; Reynolds et al 2005; Addah et al 2007).
Adolescent pregnancy complicates the already complex protein metabolism, in which protein is required for increased maternal blood volume and tissue growth, and placental and foetal growth. This makes the provision of additional dietary protein essential for the establishment and maintenance of successful pregnancy (Robinson et al 1999; O`Doherty and Crosby 1998). However, the protein requirements of the pregnant ewe also fluctuate among the different stages of pregnancy; Lippert et al (1983) have concluded that protein requirements of the ewe may be more important in mid pregnancy through its effects on lamb birth weight, than in early and late pregnancies. Conversely, similar studies have shown that protein requirements in late pregnancy are 33% higher than in mid pregnancy (Schloesser et al 1993) while in some studies, the imposition of nutritional stress in the early stages of pregnancy did not affect ovine foetal growth (Nordby et al 1987). Total nutrient intake throughout gestation may not therefore be as important as specific nutrient requirements of each stage of pregnancy. The intra-uterine growth restriction (IUGR) model has been studied and reviewed widely in many temperate sheep models (Wallace et al 1996; 1997; 2005; Reynolds et al 2005) and other species (Atinmo et al 1974; Pond et al 1991; Frank and Jane, 2006) but there is paucity of, or no literature on such studies in the adolescent Djallonké ewe. Earlier studies by Addah et al (2007) have only shown that moderate levels of under nutrition in mid pregnancy were useful in improving lamb birth weight of Djallonké ewes but the effects of dietary restrictions during the different stages of pregnancy on total and differential plasma protein, placental and birth weights of the adolescent Djallonké ewe have not been fully elucidated. This study investigated the effects of restricting dietary supplementation in early, mid or late pregnancy, on placental and birth weights of adolescent Djallonké ewes.
Twenty five (25) adolescent Djallonké(West African Dwarf) ewes were mated and used for the study at the Cattle Breeding Station of the Kwame Nkrumah University of Science and Technology, Ghana (06o43' N, 01o 36' W). A single ram of proven fertility was introduced into an adolescent female flock every morning and evening to identify oestrous females and mate them. Following a successful mating, the ewe was randomly assigned to one of five treatments in a completely randomized design and feeding of the supplementary diet and data collection commenced thence. The animals were mated between July and September 2003 and spontaneous lambing occurred between December 2003 and March 2004. The initial mean weights were 16.3 kg (T1), 15.3 kg (T2), 14.7 kg (T3), 15.1 kg (T4) and 15.4 kg (T5).
The treatments were as follows:
T1: Supplementary concentrate withdrawn in early pregnancy
T2: Supplementary concentrate withdrawn in mid pregnancy
T3: Supplementary concentrate withdrawn in late pregnancy
T4: supplementary concentrate offered throughout pregnancy
T5: supplementary concentrate completely restricted throughout pregnancy
All ewes were housed in individual pens (2.5 x 1.5 m) under natural conditions and offered a daily basal diet of 1,920 g (DM) Brachiaria decumbens plus 445.6g (DM) Gliricidia sepium hanged on top of the pens to avoid contamination while in stages of pregnancy where there were no dietary restrictions, all ewes received an additional daily concentrate of 335.1g (DM) composed of 89.1 g (DM) of whole cottonseed and 246 g (DM) of sun-dried cassava peels. The proximate compositions of both diets were determined by methods described by the AOAC (1990). The basal and supplementary diets were formulated to supply 9 and 13 ME (MJ/kg DM) and a crude protein of 6% and 12% respectively. Water was also supplied ad libitum every morning. One animal in T2 was later detected not to have conceived after two successive matings while another from T3 spontaneously aborted during late pregnancy (abortus = 400g) and were both excluded from the study. Lamb birth and placental weights were recorded within 12 h of parturition after cleaning the latter of any debris and draining it of amniotic fluids.
A total of 224 blood samples were collected for biochemical analysis. Each sample (4.5 ml) was collected by jugular venipuncture from the ewe into ethylenediamine tetra acetic acid (EDTA) impregnated vacutainers between 0800 and 0900 GMT before the provision of the rations. The initial blood samples were taken immediately after mating was successfully completed. Thereafter, samples were collected fortnightly for a period of six weeks; 3 times in each stage of pregnancy. A fourth sample was taken on the 49±1th day of each stage of gestation. Each of the three stages of pregnancy consisted of 49±1days (gestation assumed to be 147±≤3 days). The blood samples were immediately centrifuged at 3500 rpm to obtain plasma which was stored at -20oC until analysed for total protein (Tp), albumen (ALb) and globulins (GLob) concentrations by methods described by O’Doherty and Crosby (1998) and Addah et al ( 2007) with kits (PLASMATEC®) supplied by Plasmatec Laboratory Products Ltd., UK. Erythrocyte fragility due to toxicity of gossypol and HCN was assessed by visual appraisal and haemolysis was considered to have occurred if plasma exhibited a reddish colouration as opposed to the normal transparent straw-colour (Gray et al 1993).
The data collected were subjected to Analysis of Variance (ANOVA) applicable to completely randomized design using Genstat 7.1 (Lawes Agricultural Trust 2003). The effects of the various feeding regimes on the concentration of Tp, ALb and GLob, and the relationship between these metabolites, placental weight and lamb birth weight were also determined by simple linear correlation and regression analysis.
Regardless of the type of treatment, dry matter intake was highest in mid (695 g/d), lower in late (687 g/d) and lowest in early (662 g/d) pregnancy (P<0.05). The imposition of dietary restrictions in early (T1), mid (T2) or late (T3) pregnancy did not also affect the ultimate total dry matter intake of ewes in the respective treatments (P>0.05). Substantial increase in dry matter intake during late gestation is usually unattainable because of reduced volume of the abdominal cavity due to rapid increase in foetal size and associated gestational structures, which limit the distension of the rumen in the final phase of pregnancy. Reports of increased dry matter intake during mid pregnancy followed by a decline in the last one-third of pregnancy have similarly been observed in Djallonké ewes (Akinsoyinu 1985). It has also been shown that sheep offered whole cottonseed-based diets often consume relatively smaller quantities initially but ingestion is stepped up when the animals have had time to adapt (Charray et al 1992). This could explain the lower dry matter intake in the early stages of pregnancy since the feed was not initially offered to the ewes for adaptation in the pre-experimental period. The stress associated with imminence of lambing in the last few days of gestation has also been associated with reduced dry matter intake during late pregnancy (O`Doherty and Crosby 1998). Total dry matter intake in T4 was however similar to T3 but higher than T1 and T2, hence the continuous supply of the supplement throughout gestation in T4 did not increase total dry matter intake in that treatment above T3 (P>0.05). The reduction in total dry matter intake in the last phase of gestation was further demonstrated by the similarity in total dry matter intake in late pregnancy between T4, that was supplemented throughout gestation and T3, that was not supplemented in the final phase of pregnancy (P<0.05). Non-supplemented ewes (T5) however had significantly lower total dry matter intake than the other treatments (P<0.05).
No sign of erythrocyte fragility or depressed feed intake was observed. Even though the gossypol content of the concentrate diet was not determined in this study, extremely high doses of free dietary gossypol have been found to have no adverse effects on pregnancy rates and fertility in general (Gray et al 1993). Table 1 shows dry matter intake, total and differential plasma protein concentrations, and mean weight gains during pregnancy.
|
Table 1. Mean gestation, total and differential plasma protein concentrations and weight gains during pregnancy |
||||||
|
Treatment |
Gestation, Days |
dry matter intake, g/d |
Plasma Tp and differential Tp concentration, g/L |
Weight gains, kg |
||
|
Tp |
ALb |
Glob |
||||
|
T1 |
148.0 |
655.3a |
63.6a |
35.5ab |
28.5c |
4.5ab |
|
T2 |
149.8 |
673.6ab |
63.5a |
36.7b |
26.4c |
6.1a |
|
T3 |
146.7 |
646.8a |
60.9ab |
42.4c |
18.8a |
4.5ab |
|
T4 |
147.8 |
705.6b |
59.4ab |
40.7c |
18.1a |
9.6c |
|
T5 |
148.8 |
573.7c |
56.5c |
33.4a |
23.1b |
2.5b |
|
s.e.d |
0.36 |
25.66 |
1.44 |
1.55 |
2.06 |
1.07 |
|
Significance |
Ns |
* |
* |
* |
* |
** |
|
Means with different superscripts within the same column are significantly different. s.e.d = standard error of difference of means; * = P< 0.05; ** = P<0.001; Ns = not significance |
||||||
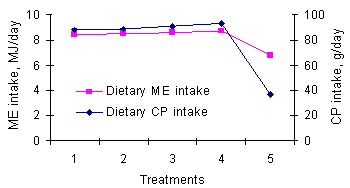
As expected, the provision of the supplementary concentrate in two (T1, T2 and T3) or three (T4) stages of pregnancy increased the dietary ME and CP in those treatments above T5 (P<0.05) but ewes receiving the supplement throughout gestation (T4) did not have higher dietary ME and CP intakes above those receiving it in only two phases of pregnancy (T1, T2 and T3) (P>0.05). The trends of dietary CP and ME intakes as illustrated in Fig. 1, were also found to be similar.
|
|
|
|
The digestive physiology of ruminants requires that dietary CP intake is synchronised with ME intake to promote efficient microbial and maternal cellular function since microbial protein synthesis is an energy-demanding process and a high or low CP intake is usually expected to induce a similar trend of ME intake (Matenga and Madsen 1992). Even though Lippert et al (1983) had earlier concluded that protein was more important than ME in alleviating the adverse effects of under nutrition during mid gestation on lamb birth weight, dietary CP intake in mid gestation for ewes in T2 ( 43.94 g/d) was lower than T1(64.8 g/d), T3(66.4 g/d) and T4 (61.0 g/d) (P<0.05) and similar to T5 (43.9 g/d) (P<0.05) yet they dropped lambs whose birth weight was similar to T1, T3 and T4 but higher than T5 (P>0.05). This is probably because microbial protein, which is a major source of protein for ruminants, is greatly influenced by energy intake than by protein intake (Robinson et al 1999) because supplemental protein sources contribute only a small proportion of the total dietary CP that escapes ruminal fermentation(Schloesser et al 1993) and may become a limiting factor for production only when ruminal amino acid supply is insufficient to meet metabolic amino acid requirements for maintenance and growth. Other findings similarly suggest that restriction of protein supplementation has no effect on lamb birth weight except when such restrictions are imposed throughout pregnancy or are chronic in nature (O`Doherty and Crosby 1998).
Total and differential plasma protein concentrations
Tp concentration was generally higher in treatments in which the concentrate supplement was withdrawn in one stage of pregnancy than those in which it was withdrawn throughout pregnancy (P<0.05). The mean plasma Tp concentration in T4 was higher than that of T5 (P<0.05), lower than those of T1 and T2 (P<0.05) but similar to that of T3 (P<0.05). It could therefore imply that withdrawing the supplement in the last phase of gestation produces similar effects on plasma TP as offering it throughout gestation. Similarly, restricting the intake of the supplement in the early (T1), mid (T2) or late (T3) stages of pregnancy did not significantly influence the mean plasma Tp concentration in the respective treatments (P>0.05). The effect only became significant when the restriction was extended to full gestation (T5) (P<0.05). This may also further suggest that maternal Tp can become a limiting plasma metabolite in pregnancy only when no supplementary diet is offered in at least two stages of pregnancy. The complete restriction of the intake of the supplementary diet throughout gestation in T5 did not significantly reduce the plasma ALb concentration in that treatment above T1 neither was the ALb and GLob concentration different between T4 and T3 (P>0.05). The effect of the different dietary regimes on Glob and ALb concentrations in T1 and T2, and T3 and T4 was also statistically insignificant (P>0.05) but Glob and ALb concentrations in T5 significantly differed from those of T2, T3 and T4 (P<0.05). In other studies, protein supplementation with soya bean meal did not affect serum Tp in any of the three stages of pregnancy but affected ALb concentration in early and mid but not late pregnancy (Schloesser et al 1993). Maternal carcass protein can only serve as a major source of protein for supporting visceral organ metabolism and foetal growth without significant effects on maternal carcass mass under moderate levels of under nutrition but not under chronic nutritional conditions (Robinson et al 1999) as imposed on T5. It therefore appears that, in sheep, Tp concentration is probably greatly influenced by the energy status of the ewe rather than by dietary CP intake since withdrawing the concentrate supplement in early (T1), mid (T2) or late (T3) pregnancy did not influence the mean Tp concentrations of those treatments compared to T4 (P>0.05). The general relationship between Tp and birth weight was however nearly linear (r = 0.93 P<0.05) while ewes with a Tp concentration threshold of ≥63 g/L (T1 and T2) gave birth to lambs with a mean birth weight of ≥1.75 kg. The highest plateau of birth weight for the Djallonké genotype has been pegged at 1.8 kg (Adu and Olaloku, 1979)
Placental, birth and gestational weights
Even though the length of gestation was not affected by the regime of dietary restrictions in the present study, limitations in placental nutrient transfer have accelerated the maturation of the foetal hypothalamic-pituitary-adrenal axis and caused early parturition in other ovine genotypes elsewhere (Wallace et al 2004). The mean weight gain of T4 was significantly greater than the other treatments (P<0.01), however no significant difference in weight gain was detected among T1, T3 and T5 (P>0.05). Based on their gestational weight gains, the ewes could be categorized into rapid (T4), normal (T1, T2 and T3) and retarded (T5) growing ewes (Addah et al 2007). The relationship between placental and lamb birth weights is illustrated by Figures 2 and 3.
|
|
|
|
|
|
The placental weights of ewes in T1 to T4 were statistically similar, though those in T2 were in absolute terms, and 38%, 21% and 24% heavier than those in T1, T3 and T4 respectively (P>0.05). The placental weights of ewes in T5 were however significantly lower than those of the other treatments (P<0.05). The birth weights in T1 to T5 were 1.67 kg, 1.84 kg, 1.51 kg, 1.54 kg and 1.17 kg respectively. The placental and lamb birth weights in T5 were significantly lower than those of the other treatments (P<0.05). Similarly, placental weights in T1 to T4 were statistically alike (P>0.05) but higher than those of T5 (P<0.05).
Reconciling the higher gestational
weight gains and yet relatively smaller placental and lamb birth weights in T4
with the relatively smaller gestational weight gains and yet higher
placental and birth weights in T2, Wallace et al (1996) and Addah et
al (2007) explained that in rapidly growing ewes, as in T4, there is
restricted transport of amino acids and glucose to the foetus and reduced
placental lactogen secretion resulting in a decreased demand on the maternal
nutrient pool, which in turn leads to increased nutrient utilization by
the maternal tissues at the expense of, and in spite of the gradually increasing
gravid uterus. The reduction in growth of the placenta of rapidly growing
adolescent ewes is associated with significant reductions in both the number of
foetal cotyledons per placenta and the mean weight of the placenta, and
reduced concentration of receptors per unit membrane due to reduced activity and
expression of system A family of amino acid and β-amino acid transporters
and/or prolonged exposure of the utero-placenatal receptors of insulin-like
growth factor 1(IGF-1) to high concentrations of foetal or maternal IGF-1 in mid
pregnancy, thereby down-regulating the placental IGF-1 expression and hence
placental growth (Wallace et al 1997; Fowden et al 2006). This ultimately
results in increased maternal growth rate and adiposity, accompanied by low
birth weight of lambs, as observed in T4. This paradoxical paradigm
of nutrient partitioning in favour of the maternal tissues and at the expense of
the gravid uterus and mammary glands appears to be unique to only adolescent
ewes in that such trajectory does not occur in overnourished primiparous adult
ewes (Wallace et al 2005). On the other hand, imposition of chronic nutritional
stress as in T5 can alter the flux of nutrients between the foetus
and its placenta to the extent that amino acids are transported from the foetus
to the placenta in order to maintain placental function, leading to foetal
wasting and subsequently lower lamb birth weight (Owens 1991). This study has
significant implications for sheep production in Africa where breeding in most
flocks is uncontrolled and adolescent/pubertal ewes with a higher potential for
growth during pregnancy get mated naturally in communal flocks at an early age
and a lower weight (Orji 1985; Karikari et al 1996). The relationship between
placental weight and a lamb birth weight as illustrated in Fig. 3 was positive
and significant(r = 0.73; P<0.05) and
is described by the regression equation:
y = 0.1x + 1.24 (R2 = 0.47)
where: y = lamb birth weight (kg); x = placental weight (g)
Hence about 47% variation in lamb birth was attributed to variation in placental weight (R2 = 0.47). Earlier results obtained by Mellor (1983) also indicate that two-thirds of the variation in birth weight of lambs was attributed to variations in placental weight. It has been further suggested that it is the small size of the placenta per se rather than alterations in its nutrient metabolism or transfer capacity that is the major limitation to foetal growth in young growing ewes (Wallace et al 2004).
O`Doherty and Crosby (1998) observed that in rapidly growing adolescent ewes, the effect of higher dietary ME intake on lamb birth weight could be smaller due to a higher efficiency of dietary ME use for maternal growth (km = 0.79) compared to conceptus growth (kc = 0.15). In another study with pregnant adolescent primiparous Djallonké ewes, those with the highest growth rates did not give birth to lambs with the highest birth weight (Adu and Olaloku 1979). The effect of rapid maternal growth during pregnancy on the reproduction of the ewe even extends to lactation where it has been proved to cause major reduction in colostrum and immunoglobulin yield compared to normal growing dams (Wallace et al 1997). The results of this study therefore suggest that nutritionally-mediated causes of low birth weight in the adolescent IUGR sheep model can be ameliorated by supplementing ewes in early (Robinson et al 1999; O’Callaghan and Bolland 1999) and late (Frutos et al 1998) pregnancy and imposing moderate levels of nutritional stress in mid pregnancy (Orr and Treacher 1994; Wilkinson and Chestnutt 1988; Addah et al 2007) when dry matter intake was highest. The advisable feeding strategy for adolescent ewes therefore is to moderately restrict or withdraw dietary supplements in mid pregnancy since this has been proved to enhance placental nutrient uptake and function, resulting in higher weights at birth
Dietary restrictions have marginal direct effects on total and differential plasma protein concentrations.
Supplementation of adolescent ewes throughout gestation impedes normal placental growth and functional capacity that leads to low birth weight while imposition of moderate levels of nutritional stress in mid pregnancy in growing adolescent ewes is recommended to improve placental function and subsequent lamb birth weight.
Addah W, Karikari P K, Osafo E L K, Oppong Anane K and Teye G A 2007 Effects of undernutrition during pregnancy on the relationship between maternal plasma metabolites concentration and lamb birth weight of adolescent Djallonké sheep. Ghanaian Journal of Animal Science 2 and 3: 184-195
Adu I F and Olaloku E A 1979 A note on nutrition during late pregnancy in West African Dwarf sheep. Animal Production 28:123-126
Akinsoyinu A O 1985 Nutrient requirements of sheep and goats in Nigeria. Proceedings of the national small ruminant production. In: Small Ruminants Production in Nigeria. (Editors Adu I F, Osinowo O A, Taiwo B B A and Alhassan W S). Proceedings of the national conference on small ruminant production held in Zaria, 6-10 October, 1985. Nigeria. pp. 141-148
AOAC (1990). Official Methods of Analysis (15th edition). Association of Official Analytical Chemists. Arlington, VA.
Atinmo T, Pond W G and Barns R H 1974 Effect of dietary energy vs. protein restriction on blood constituents and reproductive performance in swine. Journal of Nutrition 104:1033-1040
Charray J, Humbert J M and Levif J 1992 Manual of sheep production in the humid tropics of Africa. CAB International, Wallingford, UK, pp. 92-144
Fowden A L, Ward J W, Wooding F P B, Forehead A J and Constancia M 2006 Programming placental nutrient transport capacity. Journal of Physiology 572(1): 5-15. Retrieved March 28, 2008 from http://jp.physoc.org/cgi/content/full/572/1/5
Frank H B and Jane E H 2006 Fetal nutrition. In: Neonatal nutrition and metabolism (Book excerpts). (Editor William W H). 2nd edition. Cambridge University press. Retrieved March 28, 2008 from
http://www.cambridge.org/catalogue/catalogue.asp?isbn=0511189923
Frutos P, Buratovich O, Giráldez F J, Mantecón A R and Wright I A 1998 Effects of maternal and foetal traits of feeding supplement to grazing pregnant ewes. Animal Science 66:667-674
Gray M L, Greene L W and Williams G L 1993 Effects of dietary gossypol consumption on metabolic homeostasis and reproductive endocrine function in beef heifers and cows. Journal of Animal Science 71:3052-3059 http://jas.fass.org/cgi/reprint/71/11/3052
Karikari P K, Gyawu P, Asare K and Osafo E L K 1996 Assessment of the reproductive performance of a flock of Djallonké sheep bred in a humid forest environment. Journal of University of Science and Technology 16(1 and 2): 9-13
Lawes Agricultural Trust 2003 Genstats 7.1 Rothmasted Experimental Station, United Kingdom
Lippert M, Milne J A and Russel A J F 1983 Effects of mid-pregnancy supplementation on hill-ewe performance. Animal Production 36:543 (abstract).
Matenga L A and Madsen A 1992 Experiences in protein supplementary feeding of weaned lambs and goat kids in Tanzania: the issue of dietary energy. In: Small Ruminants Research and Development in Africa (Editors Rey B, Lebbie S H B and Reynolds L). Proceedings of the first biennial conference of the African Small Ruminant Research Network, ILRAD, held in Nairobi Kenya, 10-14 December 1990 by ILCA (International Livestock Centre for Africa). pp. 401-416 http://www.fao.org/Wairdocs/ILRI/x5520B/x5520b13.htm
Mellor D J 1983 Nutritional and placental determinants of foetal growth rate in sheep and consequences for the newborn lambs. British Veterinary Journal 139:307-324
Nordby D J, Field R A, Riley M L and Kercher C J 1987 Effects of maternal undernutrition during early pregnancy on growth, muscle cellularity, fibre type and carcass composition in lambs. Animal Science 64: 1419-1427
O`Callaghan D and Bolland M P 1999 Nutritional effects on ovulation, embryo development and the establishment of pregnancy in ruminants. Animal Science 68: 299-314
O’ Doherty J V and Crosby T F 1998 Blood metabolite concentrations in late pregnant ewes as indicators of nutritional status. Animal Science 66: 675-683
Orji B I 1985 Research on reproduction in small ruminants and their applicability in Nigerian production systems. In: Small Ruminants Production in Nigeria. (Editors Adu, I F, Osinowo O A, Taiwo B B A and Alhassan W S). Proceedings of the national conference on small ruminant production held in Zaria, 6-10 October, 1985. Nigeria. pp. 70 - 83
Orr R J and Treacher T T 1994 The effect of concentrate level on the intake of grass silages by ewes in late pregnancy. Animal Production 58: 109-116
Owens J A 1991 Endocrine and substrate control of fetal growth: Placental and maternal influences and insulin-like growth factors. Fertility and Development 3: 501-517
Pond W G, Maurer R R and Klindt J 1991 Fetal organ response to maternal protein deprivation during pregnancy in swine. Journal of Nutrition 121(4): 504-509
Reynolds L P, Borowicz P P, Vonnahme K A, Johnson M L Grazul-Bilska A T, Redmer D A and Caton J S 2005 Placental angiogenesis in sheep models of compromised pregnancy. Journal of Physiology 565(1): 43-58
Robinson J S, Owens J A, De Barro T, Lok F and Chidzanja M 1994 Maternal nutrition and fetal growth. In: Early fetal growth and development (Editors Wards R H T, Smith S K and Donnai D). RCOG press, London. pp. 317 – 329
Robinson J J, Sinclair K D and McEvoy T G 1999 Nutritional effects on foetal growth. Animal Science 68:315-332
Schloesser B J, Thomas V M, Petersen M K, Kott R W and Hatfield P G 1993 Effects of supplemental protein source on passage of nitrogen to the small intestine, nutritional status of pregnant ewes and wool follicle development of progeny. Journal of Animal Science 71:1019 – 1025 http://jas.fass.org/cgi/reprint/71/4/1019
Wallace J M, Aitken R P and Chayne M A 1996 Nutrient partitioning and foetal growth in rapidly growing adolescent ewes. Journal of Reproduction and Fertility 107: 183 -190
Wallace J M, Aitken R P, Milne J S and Hay W W Jnr. 2004 Nutritionally mediated placental growth restriction in the growing adolescent: Consequences for the foetus. Biology of Reproduction 71: 1055 -1062
Wallace J M, Da Silva P, Aitken R P and Cruickshank M A 1997 Maternal endocrine status in relation to pregnancy outcome in rapidly growing adolescent sheep. Journal of Endocrinology 155:359-368
Wallace J M, Milne J S and Aitken R P 2005 Effect of overnourishing singleton-bearing adult ewes on nutrient partitioning to the gravid uterus. British Journal of Nutrition 94(4):533-539
Webb R, Gosden R G, Telfer E E and Moor R M 1999 Factors affecting folliculogenesis in ruminants. Animal Science 68:285-298
Wilkinson S C and Chestnutt D M B 1988 Effects of level of food intake in mid and late pregnancy on the performance of breeding ewes. Animal Production 47: 411 -419
Received 28 April 2008; Accepted 2 June 2008; Published 4 September 2008