Genetic and non-genetic trends for calf weights in a Bos indicus herd upgraded to pedigree Brahman
D Plasse, J Arango, H Fossi, L Camaripano, G Llamozas*, A Pierre* and R Romero
Universidad Central de Venezuela, Facultad de
Ciencias Veterinarias, Maracay
*Hato Cujicito, Guardatinajas, Guarico, Venezuela
dplasse@intercable.net.ve
Abstract
(Co)variance components, genetic parameters and yearly trends of weights were estimated in a Bos indicus herd upgraded to Brahman under a technically supervised selection programme with strategic use of artificial insemination. The herd took part in a Cooperative Genetic Programme together with six others. A univariate animal model was used for birth weight (BW) and a bivariate model for weaning (205W) and 18-month (548W) weights using Restricted Maximum Likelihood. Models included random animal direct and maternal additive genetic effects, maternal permanent environmental effects and fixed effects of a year-month of birth-sex contemporary group as well as of dam age. Analysed data included 9903 BW, 9570 205W and 8393 548W of calves born in 1990 through 2001.
Estimates of direct, maternal and total heritability were for BW: 0.42, 0.07, 0.47; for P205W: 0.13, 0.14, 0.25, and for 548W: 0.22, 0.05, 0.30. Corresponding estimates of direct-maternal genetic correlations were 0.06, 0.28 and 0.34 and for the proportional permanent environmental variance due to the dam 0.01, 0.09 and 0.01, respectively. Estimates of phenotypic, direct and maternal additive genetic and non- genetic annual trends (kg) were for BW: 0.021, 0.080, -0.002, -0.062; for 205W: 2.986, 0.515, 0.418, 1.776 and for 548W: 5.773, 0.894, 0.153, 4.746.
The selection and non-genetic improvement programmes applied should be continued emphasizing cow efficiency for sustainable beef production on improved and native pasture.
Keywords: Beef cattle, Brahman, weights, genetic parameters, annual trends
Resumen
Se estimaron los componentes de (co)variancia, parámetros genéticos y tendencias anuales de pesos en un rebaño Bos indicus absorbido a Brahman y bajo un programa de selección supervisado y con uso estratégico de inseminación artificial. El rebaño formó parte de un Programa Genético Cooperativo junto con otros seis. Se usó un modelo animal univariado para peso al nacer (PN) y un modelo bivariado para peso al destete (P205) y peso a los 18 meses (P548) usando Máxima Verosimilitud Restringida. Los modelos incluyeron los efectos aleatorios aditivos directo y materno, y ambiental permanente de la vaca, como también los efectos fijos del grupo contemporáneo año-mes de nacimiento-sexo y de la edad de la madre. Los registros analizados incluyeron 9903 PN, 9570 P205 y 8393 P548 de becerros nacidos entre 1990 y 2001.
Los estimados de índices de herencia directo, materno y total fueron para PN: 0.42, 0.07, 0.47; para P205: 0.13, 0.14, 0.25 y para P548: 0.22, 0.05 y 0.30. Los estimados correspondientes de las correlaciones genéticas directo-materno fueron 0.06, 0.28 y 0.34 y para la proporción de la variancia ambiental permanente de la vaca 0.01, 0.09 y 0.01, respectivamente. Estimados de las tendencias anuales fenotípicas, genéticas aditiva directa y materna, y no genéticas (kg) fueron para PN 0.021, 0.080, -0.002, -0.062; para P205: 2.986, 0.515, 0.418, 1.776 y para P548: 5.773, 0.894, 0.153, 4.746.
Los programas de selección y mejoramiento no genético aplicados deberían continuar enfatizando la eficiencia de las vacas para una producción sostenible de carne bovina en pastos mejorados y nativos.
Palabras claves: Brahman, parámetros genéticos, pesos, tendencias anuales
Introduction
About thirty percent of Venezuelan territory is covered by savannas, which are used mainly for beef production based on a zebu population with predominantly Brahman genes. Production levels in these areas are low (Plasse 1992) although technology is available to improve them (Plasse et al 1989). Genetic improvement must be mainly through selection (Renand et al 1992). The possibility for crossbreeding with Bos taurus in extensive areas has proven to be limited by the available nutritional level and sanitary conditions (Plasse 2000), although, under certain circumstances, it might be of benefit.
Genetic researchers in Latin America depend on the collaboration of private herds to generate population statistics and prove the relative benefit of selection methods. Producers need the guidance of scientists in order to apply modern improvement technology successfully. Mutual benefit is therefore obtained in combined research-technical assistance programmes, especially where breeders´ organizations or government agencies do not fulfil the mandate of assisting the breeder. A combination of research and technical assistance makes scientific work more realistic and economic and guarantees fast application of research results in the production sector (Plasse 1981).
The present beef cattle population of Venezuela is the result of upgrading criollo cattle of Spanish origin to zebu starting in the first quarter of the last century and using bulls mainly of Brazilian and North American origin. However, the participation of the Brahman breed in the national gene pool has been predominant. The application of modern selection methods for the improvement of traits of economic importance is relatively recent in the country and is occurring only in a limited number of herds. Therefore, the objectives of the present study were to (1) obtain (co)variances and genetic parameters of a typical Bos indicus herd upgraded to Brahman and kept mainly on savanna and (2) quantify the genetic, non-genetic and phenotypic trends obtained through the application of modern selection methods based on breeding values evaluated by available appropriate statistical methods.
Materials and methods
Ranch and herd description
This research was carried out on the privately owned ranch "Cujicito" located in Guarico state, at 8° 57´ N and 67º 48´ W. The area belongs to the dry tropical forest ecosystem with a marked dry season from November through April and a rainy season from May through October. The mean annual rainfall during the years under investigation (1990 to 2001) was 1525 (1335 to 1808) mm. Three physiographic zones can be distinguished on the ranch: low, high and transition areas with temporary floods of up to 25 cm only in the low areas. Soils are acid and infertile with a mean pH of 5.5. There are nitrogen, calcium and phosphorus deficiencies. The area of native or naturalized pastures is 70% of the total and predominant species are Leersia hexandra, Panicum laxum, Hyparrhenia rufa and Dichanthium aristatum. The remaining 30% was sown during the last 15 years with introduced species, mainly Brachiaria humidicola, B. decumbens, B. arrecta and Echinochloa polystachya. This area is under strategic pasture management including fertilization and, to a limited extent, irrigation (R Tejos, personal communication).
The ranch has been the property of the same family since at least 1838. Since the 1950´s, Criollo cattle were substituted by different zebu resources by upgrading or introduction of groups of purebred zebu breeds of Cuban, Brazilian and North American origin. Between 1970 and 1985 mainly Nelore, but also Guzerat and some Indubrasil semen was used in a relatively uncontrolled upgrading system. Cows were selected for reproductive efficiency but breed characteristics dominated the selection programme. Since 1986, the herd has been included in our research-technical assistance programme and genetic, management, reproduction and sanitary programmes were designed and strictly supervised by the first author and his co-workers up to the time this paper was written.
Since 1989, only registered Brahman bulls were used in the herd, which, together with six others, was included in a Cooperative Genetic Programme (Seprocebú 2004) in that year. Between 1989 and 2000, the number of cows was reduced from 1551 to 1175, because of a decrease in land area. Since 2000, all cows are registered Brahmans and the herd functions as a Genetic Center producing registered Brahman bulls and semen for sale. Artificial insemination (AI) was introduced in the herd in the late 1950's and included up to 100% of the cows between 1970 and 1985. After 1986, strategic use of AI was practiced (Plasse et al 1988), the extent of which was limited by the restriction imposed of not sacrificing reproductive efficiency. Non-lactating cows, 2-year old heifers weighing at least 280 kg at the beginning and 320 kg in recent years, and early calving cows (more than 30 days before the breeding season) were assigned to the AI programme, using bulls of high genetic merit proven in the Cooperative Genetic Programme and young bulls on test (50% of the cows to each kind of bull). After two inseminations, or a 45-day period of AI, cows were put to selected clean-up bulls until the end of the breeding season. Apart from 73 homebred bulls used in single-sire herds and/or AI, semen of 51 sires from other herds of the Cooperative Genetic Programme and of six imported bulls was used during the years to which this study refers. In the Cooperative Genetic Programme, bulls were selected for the central semen bank on high EPDs for 18-months weight with an accuracy of at least 0.90 after having been used as young bulls in AI in at least three herds (Seprocebú 2004). Each year, two-year-old bulls to be used in natural service were selected from the respective herd on the basis of high breeding values estimated over the whole population of the Cooperative Genetic Programme using an animal model implemented with the MTDFREML programme (Boldman et al 1995). Herd sires in natural service were culled according to their EPDs for 18-months weight and not allowed to breed for more than three years. Heifers were selected for breeding at two years of age according to the target weight. However, during the transition period, until 1993, a decreasing proportion of the heifers bred were 3 years old. Non-conceiving first-service cows were eliminated and low reproductive efficiency was the main culling criterion for older cows. Reproductive problems were the principal cause of elimination (58% of all cows culled), while poor maternal ability was the cause in 14% of the losses.
The management programme was based on a limited breeding season, which was gradually shortened to 4 months during the rainy season; however, in recent years no cow was ever exposed for more than 90 days. Cows calved in calving pastures and calves were tattooed and weighed during 24 hours after birth. They stayed with their dams full time on pasture until weaning at an average of 200 days, when they were weighed again and the sexes separated. Weaned calves were kept on improved pastures (irrigated since 1994), and during the years 1997 to 2000 received a supplementation of about 0.5 kg of a mixture of cotton seed and rice meal. However, the after-weaning pasture and nutritional programmes were deficient until the year 1996. All animals had free access to a complete mineral mixture. At 18 months, young animals were weighed again. Weaning and 18-months weights were adjusted to 205 and 548 days, respectively. No selection was practiced until 18 months and all records were included in the data base.
The sanitary programme included vaccination of females against brucellosis and leptospirosis and of both sexes against foot and mouth disease, rabies and respiratory diseases. Prevention programmes against internal and external parasites were also applied. A preventive health programme for calves at birth was routine.
Statistical procedures
The original data base consisted of 10134 records of calves born alive. Of these, 231 (2.3%) records were eliminated because of birthdates outside the normal season (218), one or two progeny per sire (10), undefined breed group of dam (2) and 2-year old dam (1). The remaining 9903 calves born during the years 1990 through 2001 were offspring of 130 individual Brahman sires and a multi-sire (138 Brahman bulls) group treated as one (Table 1). They were born to 3801 dams, 79% registered Brahman cows and the rest distributed very unevenly among 11 breed group classifications: non-registered Bos indicus (7.5%), Nelore (3.5%), Guzerat (0.9%), and eight groups of different degrees of upgrading to Nelore and Guzerat (9.1%). There were 9570 data on calves weaned normally and 8393 records on 548-day weights (one birth year less).
|
Table 1. Birth years, number of records analyzed, means and standard errors for birth, 205 day and 548-day weights |
|||
|
Item |
Weights at |
||
|
birth |
205 days |
548 days |
|
|
Birth year |
1990-2001 |
1990-2001 |
1990-2000 |
|
Animals (n)a |
11799 |
|
|
|
Calves (n) |
9903b |
9570 |
8393 |
|
Sires (n)c,d |
130 |
|
|
|
Dams (n) |
3801 |
|
|
|
Progeny/sire (n)c |
61 |
|
|
|
Progeny/dam (n) |
2.6 |
|
|
|
Unadjusted means (kg) |
30.1±0.04 |
175±0.28 |
262±0.39 |
|
a
Animals for which EBVs were calculated. |
|||
To determine the relative importance of the effect of breed group of cow, preliminary analyses were carried out using general linear model techniques with the SAS programme (Freund and Littell 1981). Results from these analyses indicated that no important differences due to breed group of cow were present, so the analyses were continued excluding this effect. In order to determine the other fixed effects to be included in the model, preliminary analyses were performed for birth, 205-day and 548-day weights using mixed models with the SAS (Mixed Procedure) programme (Littell et al 1996) including sire as a random effect. Year of birth, month of birth and their interaction, sex and age of dam were highly significant for all weights; however, generation was not significant for 205-day or 548-day weights and was excluded from the final model. One pre-analysis was intended to test the importance of the random effect of the sire x year interaction. Using a likelihood ratio test, the comparison of the results from the model including or excluding this interaction showed that no improvement was obtained by including it and therefore it was not considered in the final animal model. The final animal model defined a year-month of birth-sex contemporary group and included age of dam also as a fixed effect.
A univariate model was used for the analysis of birth weights and a bivariate model for 205-day and 548-day weights.
The basic model was:
y = Xb + Za + Zm + Wc + e
where for the univariate case:
y is the
Nx1 vector of records,
X, Z, and W
are the known incidence matrices that associate levels of b, a, m and c with y,
b denotes the
unknown vector of fixed effects,
a is the
unknown vector of breeding values for direct additive genetic effects,
m is the
unknown vector of breeding values for maternal additive genetic effects,
c is the
unknown vector of permanent environmental effects contributed by dams to records
of their progeny, and
e is the
vector of residual effects.
Estimation of (co)variance components was carried out by restricted maximum likelihood employing a simplex algorithm to search for variance components to minimize the -2log Likelihood (L) (Boldman et al 1995). Convergence was assumed when the variance of the function values (2log L) of the simplex was at least 10-7. After first convergence, restarts were performed to verify that it was not a local minimum.
Breeding values were predicted utilizing all pedigree information available. Annual genetic trends were estimated for each trait by regressing the mean breeding values of calves on the corresponding birth year. Phenotypic trends were estimated by regressing the unadjusted weights on birth year. Non-genetic trends were estimated from the year solutions of the animal model analyses and express yearly differences practically free of genetic effects.
The main fixed effects included in the final animal model had the following classifications: birth year, 1990 to 2001 (for P548 1990 to 2000); month of birth, 2 to 6; sex, males, females; age of dam, 3 to 13+ years.
Results and discussion
Data structure and means
The data structure is given in Table 1. The mean number of progeny per sire for calves with known paternity was 61, higher than in Plasse et al (2002a,b) and very satisfactory. The mean number of progeny per dam was 2.6. Of all calves born, 80% were offspring produced by AI or in single-sire herds. This percentage is lower than found in Plasse et al (2002b) where it was 100%, but higher than in Plasse et al (2002a), where it was 47%. However, the proportion of calves born with known paternity increased over the years up to 98% in 2001. Where AI is used strategically in the tropics, a varying, but sometimes high proportion of cows not apt for this technique needs to be assigned directly to natural service or, if they fail to conceive to AI, to clean-up bulls. Most ranches cannot afford to make sufficient paddocks for such a large percentage of cows to be bred in single-sire herds, because of the high investment required or natural limitations, which prevent subdivision of native grassland. Therefore some cows might be bred in multi-sire herds and paternity of their calves is not known. Obviously, animals without known paternity cannot be registered and in this ranch they were sold to commercial herds or fattened.
Compared to the summaries for this breed from the Latin American literature published by Plasse (1978) and updated by Arango and Plasse (1994) and to more recent publications by Romero et al (2001) and Plasse et al (2002a,b), birth and weaning weights (Table 1) were higher than the means published, except the values reported by Romero et al (2001) for Brahman on cultivated pasture. The mean for 548-day weight was similar to those found in the literature review by Arango and Plasse (1994), but less than means reported by Romero et al (2001) and Plasse et al (2002a,b,) all from more favourable conditions.
There were only 153 inbred animals with average inbreeding of 7.7 %.
Birth weight
Estimates of (co)variance components and genetic parameters for 9903 birth weights are given in Table 2. Direct heritability was 0.42±0.04.
|
Table 2. Estimates of (co)variance components and genetic parameters for birth (BW), 205-day (205W) and 548-day (548W) weights (kg) |
||||||
|
Trait |
Variance componentsa |
|||||
|
s2a |
s2m |
sam |
s2c |
s2e |
s2p |
|
|
BW |
6.41 |
1.13 |
0.15 |
0.18 |
7.45 |
15.3 |
|
205W |
49.2 |
53.9 |
14.5 |
33.0 |
235 |
385 |
|
548W |
129 |
32.5 |
22.2 |
3.50 |
413 |
601 |
|
Trait |
Parametersb |
|||||
|
h2a |
h2m |
h2t |
ram |
c2 |
e2 |
|
|
BW |
0.42±0.04 |
0.07±0.02 |
0.47 |
0.06±0.13 |
0.01 |
0.49 |
|
205W |
0.13 |
0.14 |
0.25 |
0.28 |
0.09 |
0.61 |
|
548W |
0.22 |
0.05 |
0.30 |
0.34 |
0.01 |
0.69 |
|
|
Correlations between 205 and 548-day weightsc |
|||||
|
ra1a2 |
rm1m2 |
rc1c2 |
re1e2 |
rp1p2 |
|
|
|
|
0.85 |
0.72 |
1.00 |
0.59 |
0.66 |
|
|
a
s2a:
direct additive genetic variance;
s2m:
maternal additive genetic variance;
sam:
covariance between direct additive and maternal additive genetic effects;
s2c:
maternal permanent environmental variance;
s2e:
temporary environmental variance;
s2p:
phenotypic variance. |
||||||
This estimate is higher than the 0.23 and 0.33 reported by Plasse et al (2002a,b) from two Brahman herds in our programme, which are kept under similar selection and management schemes, but are different in original genetic composition, years of selection, ecologic condition and number of cows, which might affect the proportional variances in this and the other two weights and thus the estimated parameters. The values are also higher than the mean reported by Mercadante et al (1995) in a world literature review of Bos indicus cattle, which was 0.33. The mean for Bos indicus and Bos taurus cattle in a review by Koots et al (1994a) was 0.31. Maternal heritability was 0.07±0.02, equal or similar to the results in Plasse et al (2002a,b), while Koots et al (1994a) reported an average of 0.14 and Mercadante et al (1995) of 0.12. Total heritability (Willham 1972) was 0.47, higher than the estimates found in two other Brahman herds in Venezuela (Plasse et al 2002a,b), which were 0.30 and 0.28 respectively. The correlation between direct genetic and maternal genetic effects was 0.06±0.13, while Plasse et al (2000a,b) published estimates of 0.22 and -0.37, and Koots et al (1994a) and Mercadante et al (1995) obtained mean estimates in their reviews of -0.35 and -37, respectively. Permanent environmental effects (c2) contributed 1% to the phenotypic variance, which is lower than the 4% and 3%, respectively, reported by Plasse et al (2002a,b).
205-day weight
Estimates of (co)variances and genetic parameters for 9570 weaning (205-day) weights are reported in Table 2. The estimated direct heritability of 0.13 is much higher than those found in previous publications from two herds in our programme (Plasse at el 2002a,b), which were 0.08 and 0.07, but it is lower than the means reported in the reviews of Koots et al (1994a) and Mercadante et al (1995), of 0.24 and 0.22, respectively. Maternal heritability was 8% higher than direct heritability while Plasse et al (2002a,b) found it to be 75% and 100% greater than direct heritability but similar in absolute terms (all 0.14). On the other hand, in their review Koots et al (1994a) found maternal heritability to be, on average, 54% of direct heritability. Maternal heritability was 45% of direct heritability in Zebu females in Brazil (Mercadante y Lôbo 1997). In these two publications the absolute value for maternal heritability was 0.13, almost equal to ours. While the present estimate for maternal heritability was equal to that in our earlier papers and very similar to the means obtained in the literature reviews cited, direct heritability was higher than our earlier estimates, but lower than the means found in the cited reviews. Total heritability was 0.25, considerably higher than our earlier estimates in two Brahman herds (Plasse et al 2002a,b), which were 0.16 and 0.12, but lower than the estimate obtained by Mercadante and Lôbo (1997) which was 0.35.
The direct-maternal genetic correlation was 0.28, while our previous values published were 0.07 and -0.13 (Plasse et al 2002a,b). The estimates found in the reviews by Mercadante et al (1995) and Koots et al (1994a) were -0.23 and -0.16. Our moderate positive genetic correlation between direct and maternal effects contrasts with most published estimates. Robinson (1996a) concluded that "negative estimates were more likely to be a consequence of additional variation between sires or sire x year variation, than evidence of a true negative genetic relationship", and suggested the need for further research into this matter. When including a sire x year interaction in a full animal model, Robinson (1996b), Lee and Pollak (1997) and Meyer (1997) obtained a decrease in the absolute value of negative direct-maternal correlations. In the present analysis this interaction was not considered in the final model because its inclusion did not improve the -2log Likelihood significantly in a preliminary analysis. On the other hand, Meyer (1997) postulated that less negative correlations were obtained in experimental than in field data, which might explain our positive correlation in data from a well controlled breeding programme. While Mercadante and Lôbo (1997) reported a mean estimate of 11% from the literature and 10% from a Nelore herd in Brazil, for the contribution of the permanent environmental variance to the phenotypic variance, Plasse et al (2002a,b) obtained estimates of 14% and 16%. The present estimate, 9%, is lower.
548-day weight
For 8393 weights at 548 days direct, maternal and total heritabilities were 0.22, 0.05 and 0.30, all being similar or higher than found in our previous publications from two Brahman herds in Venezuela, where they were 0.16, 0.04, 0.28 (Plasse et al 2002a) and 0.13, 0.08, 0.26 (Plasse et al 2002b), respectively. The c2-effects (1%) were the same as in Plasse et al (2002b), but much lower than in Plasse et al (2002a) in which they accounted for 4% of the total variance. The direct-maternal correlation of 0.34 contrasts with the corresponding estimates in Plasse et al (2002a,b), which were 0.86 and 0.49.
Most papers published on post-weaning weights have been carried out with yearling weights and the results are not comparable with our data on 18-months weight, because in the tropics the after-weaning stress period usually extends beyond one year of age. For this reason, we consider that under our conditions 18-months weight is more appropriate for assessing the genetic differences in post-weaning weights and for use in the selection programme.
Correlations between 205- and 548-day weights
The direct genetic correlation between 205-day and 548-day weight was 0.85 (Table 3), higher than the estimates of 0.66 and 0.64 found in two other Brahman herds in Venezuela (Plasse et al 2002a,b), and the mean for zebu cattle summarized by Mercadante et al (1995), which was 0.71. It was similar to the mean of 0.81 calculated by Koots et al (1994b) for the genetic correlation between weaning and yearling weight in a world literature review, while Meyer (1994) found a genetic correlation of 0.69 between weaning and final (16 - 23 months) weights from crossbred Australian zebus.
The maternal genetic correlation was 0.72, while the estimates in our previous publications were 0.70 and 0.96 respectively (Plasse et al 2002a,b). The permanent environmental correlation was unity as well in Plasse (2002a,b). From the high maternal genetic and permanent environmental correlations between weaning and 18-months weight, it can be concluded that important genetic and permanent environmental effects due to the dam might still be present at 18 months in tropical beef cattle. This can be explained as a carry-over effect from the pre-weaning period. Such observations have been made by other authors (Meyer 1994; Tosh et al 1999; Plasse et al 2002a,b). The estimate of the residual correlation of 0.59 was very similar to that published by Plasse et al (2002a) but lower than the estimates given by most other authors between weaning and 12-months and between weaning and 18-months weight (Meyer 1994; Eler et al 1995; Plasse et al 2002b) for tropical beef cattle. The phenotypic correlation of 0.66 between 205 and 548-day weight was similar to estimates summarized by Mercadante et al (1995) from the world literature on zebu cattle, which was 0.64, as was also the estimate of Plasse et al (2002a) for another Brahman herd. Plasse et al (2002b) found in a third Brahman herd a higher phenotypic correlation of 0.74.
Our results on heritability and genetic correlation estimates indicate that milk production in this herd could be improved by selecting for maternal effects at weaning and that this would not be antagonistic to genetic improvement of growth rate. Growth rate should be improved by selection for 18-months weight rather than using weaning weight as a criterion. Such a procedure would also guarantee a simultaneous improvement of milk production in the population, because of the high genetic correlation between maternal effects of 18-months and weaning weights. However, we understand that selection for reproductive efficiency is primary and improvement of milk production and growth is secondary and limited by a sustainable pasture improvement program.
Phenotypic trends
The annual phenotypic trend for birth weight was irregular and non-significant and the mean close to zero (Figure 1a). However, the mean of 30 kg for birth weight (Table 1) is satisfactory and phenotypic improvement of this trait in the herd is not essential, specially since pre-weaning mortality during the respective years was on average 2.6% (unpublished), which is very low for tropical beef cattle. For 205-day weight, the mean annual phenotypic trend was 2.99 kg (Figure 1b) and quite linear, highly significant and similar to values reported by Hoogesteijn and Verde (1998) and Plasse et al (2002a) in two other Brahman herds in Venezuela. However, it was higher than the mean trend obtained by Plasse et al (2002b) in a third Brahman herd, which was 0.471 kg. From 162 kg in the first year (1990), weaning weight increased to 193 kg in 2001 after having reached 203 kg in the previous year. Mean 548-day weight increased from 220 kg in the first year to 294 kg in the last year (Figure 1c) generating a highly significant mean trend of 5.77 kg per year, much higher than the means of 1.5 kg (Hoogesteijn y Verde 1998), 1.8 kg (Plasse et al 2002a) and 2.0 kg (Plasse et al 2002b) reported from three other Brahman herds in Venezuela.

Figure 1a.
Annual
phenotypic trend (kg) by year of birth for birth weight (BW)
nsNon-significant

Figure 1b.
Annual
phenotypic trend (kg) by year of birth for 205-day weight (205W)
++P<0.01

Figure 1c.
Annual
phenotypic trend (kg) by year of birth for 548-day weight (548W).
++P<0.01
Direct additive genetic trends
The estimate for annual direct additive genetic trend for birth weight was 0.080 kg, highly significant and quite linear (Figure 2a). It is apparently a correlated response to selection for 548-day weight in bulls and maternal ability in cows. It is similar to the 0.061 kg obtained by Plasse et al (2002b), while Plasse et al (2002a) did not observe any significant change. For 205-day weaning weight, the estimate for annual direct additive genetic trend was 0.515 kg (Figure 2b), highly significant and quite linear. This estimate is higher than those obtained by Plasse et al (2002a,b) which were 0.142 and 0.126 kg, probably because of higher selection pressure on cows due to an important decrease in herd size during the years 1989 through 1999. The estimate of annual direct additive genetic trend for 548-day weight was 0.894 kg, very linear (Figure 2c) and highly significant. This estimate was higher than those obtained by Plasse et al (2002a,b) which were 0.263 and 0.486 kg respectively, covering longer time periods. Another reason is that this is a large herd with a relatively high percentage of cows in artificial insemination (65% during the corresponding years) and it makes efficient use of the best bulls of the semen bank of the Cooperative Genetic Programme to which it belongs. Compared to the other two herds mentioned, which participate in the same programme, it also has higher estimates for direct heritability for 548-day weight. Mean direct additive genetic trend per year in the whole population of the cooperative was 1.4 kg during the years 1990 through 2002 (Seprocebú 2003).
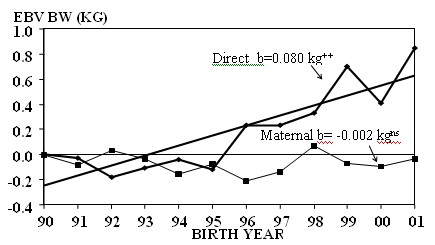
Figure 2a.
Annual direct and maternal genetic trends (kg) by year of birth for birth weight
(BW);
EBV, estimated breeding value (regression lines are for direct trends)
nsNon-significant:++P<
0.01

Figure 2b. Annual direct and maternal genetic trends (kg) by year of birth for 205-day weight (205W) ;
EBV, estimated breeding value (regression lines are for direct trends)
++P<
0.01

Figure 2c.
Annual direct and maternal genetic trends (kg) by year of birth for
548-day weight (548W);
EBV, estimated breeding value (regression lines are for direct trends)
++P<
0.01
Since the oldest cows in the data set were born in 1977, breeding values were also available for animals born between then and 1989. However, the breeding values of these animals were predicted without their own records, which were not available and the calculations were based on the weights of their descendants. Therefore the accuracies of their breeding values would be low. Direct genetic trends were 0.166 and 0.894 kg for the time periods 1977 through 1989 and 1990 through 2001 respectively, showing that the genetic trend was very low in the time period where little or no selection was practiced for weights, while it was considerable after 1990, when bulls with higher breeding values and greater accuracies, mainly for 18-months weight, were used, and the herd took part in a Cooperative Genetic Programme.
Mean maternal additive genetic trend for birth weight was non-significant (Figure 2a) while for 205-day and 548-day weights estimates were 0.418 and 0.153 kg and highly significant. The trend for the two latter weights was quite linear (Figures 2b and 2c). Maternal additive genetic trend for weaning weight reflects the mean annual genetic change in milk production, and shows that annual lactation milk yield increased enough to support an increase in weaning weight of 0.418 kg per year (Figure 2b). This maternal additive genetic trend at weaning was much higher than the 0.115 and 0.044 kg in two other Brahman herds of our programme (Plasse et al 2002a,b), probably due to higher selection for maternal ability. While for those two herds maternal additive genetic trends for 548-day weight were 0.095 and 0.251 kg, in the present analysis it was 0.153 kg and highly significant (Figure 2c). Maternal additive genetic trend at 18 months has to be seen as a carryover effect from the genetic improvement of milk production and growth potential in the pre-weaning period. Direct and maternal additive genetic trends are expressed phenotypically in different generations. However, over a longer time period it should be acceptable to add both means in order to obtain an estimate for total genetic trend achieved for a particular weight by direct and correlated response through selection for growth rate to 18 months in males and maternal ability in females. For 205-day and 548-day weights in this herd, this would amount to 0.933 and 1.047 kg per year. This would mean that of the phenotypic change in 12 years of 36 kg for 205-day weight and 69 kg for 548-day weight, 11 and 13 kg respectively, would have been due to total genetic additive improvement during the 12-year period. The total additive genetic trend is, in part, a consequence of a reasonably high correlation between direct and maternal additive genetic effects for each of these two weights and high direct and maternal additive genetic correlations between them. The important contribution of maternal genetic to total genetic trend for weaning weights permits the conclusion that cow selection for genetic improvement of weaning weight was effective. As a reference for comparison with genetic change in a large cooperative genetic programme, Crump et al (1997) found the mean annual genetic trend for 400-day weight of the registered populations of Simmental, Limousin, Charolais, South Devon and Angus breeds in Great Britain to vary from 1.04 to 2.49 and 0.82 to 1.86 kg, respectively, for the period 1980 to 1992 and for all years on record.
Non-genetic trends
Estimates of non-genetic trends for 205-day and 548-day weights calculated from the year solutions had means of 1.78 (non-significant) and 4.75 kg (highly significant) for weights at 205 and 548 days of age. The linear trend for birth weight was close to zero and non-significant. In the latter, the drop between 1991 and 1994 (Figure 3a) was an expected consequence of a leptospira infection of the herd and accompanied by an increase in the abortion rate. The solutions also showed a depression between 1995/94 and 1998/97 for 205-day/548-day weights (Figures 3b and 3c), which correspond to birth years with a deterioration of pasture quality and quantity. In the case of after-weaning management, this was later corrected with supplementation and/or the use of irrigated cultivated pastures. The high mean non-genetic trend for 18-months weight demonstrates the result of successful after-weaning environmental improvement programmes, such as pasture management, sanitary control and supplementation.

Figure 3a.
Annual
non-genetic trends (kg) by year of birth for birth weight (BW). SNG, solutions
for years.
nsNon-significant

Figure 3b.
Annual
non-genetic trends (kg) by year of birth for 205-day weight (205W). SNG,
solutions for years.
nsNon-significant

Figure 3c.
Annual
non-genetic trends (kg) by year of birth for 548-day weight (548W). SNG,
solutions for years.
++P<0.01
Conclusions
- The results confirm our opinion that artificial
insemination can be successfully employed in a large tropical beef
cattle herd and may contribute to genetic and phenotypic progress.
However, the programme must be reasonably designed, and its
application matched to the stage of technical development of the
herd, the intensity of the management programme and the
infrastructure of the ranch. To be really successful, it also
needs to be accompanied by sufficient pasture supply, strategic
supplementation and sanitary control, so that cattle are able to
express their genetic growth and reproductive potential in terms of
better phenotypic production.
- Although the initial weights were low, but comparable
to most other values published for the Latin American Tropics,
after 12 years of genetic and non-genetic improvement programmes,
they reached very satisfactory yearly means. Annual phenotypic and
genetic trends were higher than in two other herds analyzed from
our programme. Genetic trend for milk production potential was also
evident. The non-genetic change at weaning may be seen mainly as
the consequence of better pasture supply to the cow herd and a more
intensive sanitary programme for calves. The high non-genetic trend
for 548-day weight was due to an improvement of pasture management,
sanitary programme and supplementation in the after-weaning period.
Along with the improvement of weights, pre- and post-weaning
mortality was decreased and pregnancy percentage tended to increase
although still fluctuating between years (non-published
data).
- Estimates of direct and maternal heritabilities were
higher or similar to those reported in the corresponding
literature. Those of direct-maternal genetic correlations were
positive for the three weights, confirming again that in a well
designed breeding and selection programme apparent negative
estimates for this correlation may be avoided. The sire x year
interaction was not significant.
- The selection programme used in this herd emphasized reproductive efficiency in cows and growth rate up to 18 months, estimated by 548-day weight, in sires. The genetic and non-genetic programmes have been successful and in the future should emphasize cow efficiency for improved and sustainable beef production on natural and strategically improved pastures.
Acknowledgments
The authors thank Florangel Quero and Mayanin Dagger for assistance in the preparation of data for the analyses. The authors most gratefully acknowledge the contribution of Professor Lucia Vaccaro in the preparation of the manuscript. The analyses of the data were supported by the Consejo de Desarrollo Científico y Humanístico (UCV) through Proyecto de Grupo Nº 11-10-4981-02 and Ayuda Institucional "A" Nº 11.10-5027-2002, which is gratefully acknowledged.
References
Arango J and Plasse D 1994 Crecimiento en cruces de razas cebuinas. In: X Cursillo sobre Bovinos de Carne. Editors: D Plasse, N Peña de Borsotti, J Arango. Universidad Central de Venezuela, Facultad de Ciencias Veterinarias, Maracay, Venezuela, pp 159-198.
Boldman K G, Kriese L A, Van Vleck L D, Van Tassell C P and Kachman S D 1995 A Manual for Use of MTDFREML. A Set of Programs to Obtain Estimates of Variances and Covariances. US Department of Agriculture, Agricultural Research Service, Clay Center, NE, USA. 114 pp.
Crump R E, Simm G, Nicholson D, Findlay R H, Bryan J G E and Thompson R 1997 Results of multivariate individual animal model genetic evaluations of British pedigree beef cattle. Animal Science 65: 199-207.
Eler J P, Van Vleck L D, Ferraz J B S and Lôbo R B 1995 Estimation of variances due to direct and maternal effects for growth traits of Nelore cattle. Journal of Animal Science 73:3253-3258.
Freund R and Littell R 1981 SAS® for Linear Models. A Guide to the ANOVA and GLM Procedure. SAS® series in Statistical Applications, SAS Institute Inc., Cary, NC., USA. 231 pp.
Hoogesteijn R y Verde O 1998 Implementación y resultados de un programa de mejoramiento productivo en un rebaño Brahman registrado en el estado Apure. In: XIV Cursillo sobre Bovinos de Carne. Editors: D Plasse, N Peña de Borsotti, R Romero. Universidad Central de Venezuela, Facultad de Ciencias Veterinarias, Maracay, Venezuela, pp 167-214.
Koots K R, Gibson J P, Smith C and Wilton J W 1994a Analyses of published genetic parameter estimates for beef production traits. 1. Heritability. Animal Breeding Abstracts 62:309-338.
Koots K R, Gibson J P and Wilton J W 1994b Analyses of published genetic parameter estimates for beef production traits. 2. Phenotypic and genetic correlations. Animal Breeding Abstracts 62:825-853.
Lee C and Pollak E J 1997 Relationship between sire x year interactions and direct-maternal genetic correlation for weaning weight of Simmental cattle. Journal Animal Science 75:68-75.
Littell R, Milliken G A, Stroup W W and R Wolfinger W 1996 SAR® System For Mixed Models. SAS® Institute Inc. Cary, NC, USA. 633 pp.
Mercadante M E Z y Lôbo R B 1997 Estimativa de (co)variãncias e parâmetros genéticos dos efeitos direto e materno de características de crescimento de fêmeas de um rebanho Nelore. Revista Brasileira de Zootecnia 26: 1124-1133.
Mercadante M E Z, Lôbo R B y Borges A R 1995 Parámetros genéticos para características de crecimiento en cebuinos de carne. Archivos Latinoamericanos de Producción Animal 3: 45-69.
Meyer K 1994 Estimates of direct and maternal correlations among growth traits in Australian beef cattle. Livestock Production Science 38: 91-105.
Meyer K 1997 Estimates of genetic parameters for weaning weight of beef cattle accounting for direct-maternal environmental covariances. Livestock Production Science 52: 187-199.
Plasse D 1978 Aspectos de crecimiento del Bos indicus en el Trópico Americano (Primera parte). World Review of Animal Production XIV, 4:29-48.
Plasse D 1981 Integración de investigación y asistencia técnica para disminuir la dependencia alimenticia de América Latina. Asociación Latinoamericana de Producción Animal, Memorias 16:21-26.
Plasse D 1992 Presente y futuro de la producción bovina en Venezuela. In: Ganadería Mestiza de Doble Propósito. Editor: C Gonzalez-Stagnaro. Universidad de Zulia, Facultad de Agronomía y Ciencias Veterinarias, FUSAGRI, GIRARZ. Maracaibo, Venezuela. pp 1-24.
Plasse D 2000 Cruzamiento en bovinos de carne en América Latina Tropical: Que sabemos y que nos falta saber. In: III Simposio Nacional de Melhoramento Animal. Editor: I. J.Nunes, F. E. Madalena, M. de A. E. Silva. Sociedade Brasileira del Melhoramento Animal, Belo Horizonte, Minas Gerais, Brasil. pp 165-179.
Plasse D, Hoogesteijn R, Fossi H, Verde O, Bastidas P, Rodríguez R and Silva V 1988 Estrategias para el uso de la Inseminación Artificial en Bovinos de Carne en Venezuela. Universidad Central de Venezuela, Facultad de Ciencias Veterinarias. Maracay, Venezuela. 119 pp.
Plasse D, Linares T, Hoogesteijn R and Fossi H 1989 Programa de Aplicación Tecnológica para Mejoramiento de la Producción Bovina de Carne. Facultad de Ciencias Veterinarias-UCV y Programa de Cooperación Agrícola SPB MIG: Convenio MAC-PDVSA, Boletín No. 1, Caracas, Venezuela. 50 pp.
Plasse D, Verde O, Arango J, Camaripano L, Fossi H, Romero R, Rodríguez C and Rumbos J 2002a (Co) variance components, genetic parameters and annual trends for calf weights in a Brahman herd kept on floodable savanna. Genetics and Molecular Research 1 (4): 282-297. http://www.funpecrp.com.br/gmr/year2002/vol4-1/gmr0044_full_text.htm. (accessed January 30, 2004).
Plasse D, Verde O, Fossi H, Romero R, Hoogesteijn R, Bastidas P and Bastardo J 2002b (Co)variance components, genetic parameters and annual trends for calf weights in a pedigree Brahman herd under selection for three decades. Journal of Animal Breeding and Genetics 119(3): 141-153.
Renand G, Plasse D and Anderson B. B. 1992 Genetic improvement of beef cattle for growth and carcass traits. In: Beef Cattle Production. World Animal Science, C5. Editors: R Jarrige and C Béranger. Elsevier, Amsterdam, Netherlands. pp 87-104.
Robinson D L 1996a Estimation and interpretation of direct and maternal genetic parameters for weights of Australian Angus cattle. Livestock Production Science 45: 1-11.
Robinson D L 1996b Models which might explain negative correlations between direct and maternal genetic effects. Livestock Production Science 45: 111-122.
Romero R, Plasse D, Verde O, Hoogesteijn R, Bastidas P and Rodríguez R 2001 Absorción de Brahman a Guzerá y Nelore en pasto mejorado 2. Pesos al nacer, destete y dieciocho meses. Livestock Research for Rural Development 13 (1). Available at http://www.cipav.org.co/lrrd/lrrd13/1/rome131.htm (accessed July 30, 2003).
SEPROCEBÚ 2004 Sementales Probados Cebú, C.A, 2004. Available at www.seprocebu.com/genetico.htm (accessed March 15, 2004).
Tosh J J, Kemp R A and Ward D R 1999 Estimates of direct and maternal genetic parameters for weight traits and backfat thickness in a multibreed population of beef cattle. Canadian Journal of Animal Science 79: 433-439.
Willham R L 1972 The role of maternal effects in animal breeding: III-Biometrical aspects of maternal effects in animals. Journal of Animal Science. 35: 1288-1302.
Received 13 February 2004; Accepted 4 May 2004