Genotype by environment interaction for milk production traits in Iranian Holstein dairy cattle using random regression model
Mehdi Bohlouli and Sadegh Alijani
Department of Animal Science, University of Tabriz, Tabriz, Iran
M.bohluly@gmail.com
Abstract
In this research the existence of genotype by environment interaction (G × E)on milk production traits (milk yield, fat yield, protein yield, fat percentage and protein percentage) was investigated by considering relevant records in different production levels as different traits. Production level (PL) considered as the average milk production traits of herd-year calving.Data on Holstein cows from 2001 to 2010 were used. Herd-years of calving means for milk production traits were clustered in three levels using the FASTCLUS procedure in SAS software. Production levels included low, medium and high levels. For these traits, G × E were investigated by applying a multiple-trait random regression sire model.
Additive genetic and permanent environmental variances of milk production traits varied in different production levels. Estimated heritabilities for milk production traits as a function of days in milk were highest in high production levels with an exception for protein percentage. Generally, the highest heritability estimates of 305-d milk production traitswere found in high PL rather than low PL. Low spearman correlations between estimated breeding values of the 20 top sires among low and medium PL (0.38) and between low and high PL (0.39) of milk yield showed re-ranking of sires for these levels.The greatest G×E was observed for milk yield and protein percentage, with a genetic correlation for 305-d equal to 0.79 between low and high production levels. Results from this research indicated that milk production of daughters of the same sires depends greatly on the production environment.
Keywords: Genetic Correlation, Genotype by Environment Interaction, Production Levels, Random Regression Model
Introduction
The environments in which dairy farming is practiced in Iran vary in many ways, such as the average herd production, level of feeding, elevation and climate variables including temperature and humidity. The phenomenon by which different genotypes respond differently to changes in their environments is known as genotype by environment interaction (G×E) or as differences in environmental sensitivity of genotypes (Falconer and Mackay 1996). This interaction can cause different ranking of animals across environments or a change of scale, i.e., variance, across environments (Lynch and Walsh 1998). If a genotype by environment interaction exists, the ranking of sires for milk production traits of their daughters will vary from one environment to another. However, re-ranking of sires across environments is limited for milk production traits (Veerkamp et al 1995; Cromie et al 1998; Calus et al 2002); although there is evidence that variances and heritabilities vary. An environmental parameter reflected the environment encountered by the animals; such as production level of herds (Veerkamp and Goddard 1998; Calus et al 2002), or other characteristics of the herds, such as average age at calving (Fikse et al 2003).
Typically, estimated genetic correlations across environments are used to estimate the degree of re-ranking. Genotype by environmental interaction is reflected by the genetic correlation between different production levels. High estimates of genetic correlations between environments (>0.80) suggest no evidence for strong G × E (Robertson 1959). Using herd production level as a substitute for the level of feeding, Veerkamp and Goddard (1998) reported a genetic correlation of 0.79 between Australian dairy herds with <20 kg of milk yield per day and herd with >24 kg of milk production per day.
Calus et al(2002) reported similar results for genetic correlations between extreme classes of herd production level in Dutch dairy cattle.Hammami et al (2008) reported that genetic correlation for 305-days milk between Tunisia and Luxemburg countries was 0.60 and spearman rank correlation between estimated breeding values (EBV) of common sires for this trait from within-country analyses was 0.41. Kolver et al (2002) reported significant G×E for milk and fertility traits when the performance of New Zealand and imported Holstein Friesian dairy cows were compared on all pasture or total mixed ration systems.The purpose of the present research was to estimate genotype by environment interaction for milk production traits and to investigate the effects of production levels on re-ranking of sires EBVs.
Material and methods
Data
Total of test-day records for first lactation Holstein cows from 2001 to 2010 were extracted from the Animal Breeding Center database at Karaj, Iran. Records were edited on the following criteria: cows with known sires and having age at first calving from 21 to 46 month, edits excluded irregular data for daily milk yield (<1.0 and >75 kg), fat percentage (<1.5% and >9%), and protein percentage (<1% and >7%), cows were required to have a minimum of five test-day (TD) records between 5 and 305 days in milk (DIM) and herd-year of calving subclasses with at last 10 cow records. Additional data edits eliminated sires that had progeny in fewer than three herds and herds that used fewer than three sires.
The FASTCLUS procedure of SAS software (SAS 2003) was used to clustering and average of trait in herd-year of calving grouped in three clusters. Common sires had at last 10 daughters in each group of production levels. The pedigree file included animal code, sire code and maternal grandsire code. Therefore, all of the sires had genetic relationship with common sires kept in the analysis.
Statistical analysis
For examining a genotype × production levels interaction, the performances of cows in production levels were considered as different traits. Thus, the following multiple-trait sire model, which considered the performance of daughters of sires in the three production levels as different correlated traits, was fitted:

The following (co)variance structure was assumed for random effects of model:

where G is sire genetic (co)variance matrix among random regression coefficients and A is additive numerator relationship matrix between sires. The matrix P was the cow effects variance-covariance matrix among random regression coefficients, and e was residual variances for each traits and I represents an identity matrix with ones on the diagonal. G and P are 12×12 (co)variances matrix of regression coefficients. All across-production levels (co)variances in P equal to zero because this effect was considered independent across production levels.
The first four Legendre polynomial functions (Kirkpatric et al 1990) were given as:

where w is a standardized unit of DIM and ranged from -1 to +1. Estimated (co)variance component of milk production traits were obtained by REMLF90 program based on restricted maximum likelihood method (Misztal et al 2002).
Calculation of genetic parameters
The
sire and permanent environmental (co)variances matrices for each DIM were
calculated as:


In sire model analysis, EBVs were computed via multiplying sire additive genetic value by 2. Spearman rank correlations between EBV of common sires in each group were used to assess the level of re-ranking of sires in different production levels for milk yield, protein yield, fat yield, protein percentage and fat percentage calculated using the procedure CORR of SAS (SAS2003).
Results and discussion
Descriptive statistics
The mean, standard deviation and coefficient of variation of milk production traits and other descriptive statistics for different production levels are summarized in Table 1. The number of cows per common sires was the lowest in the low production level and highest in high production level for milk yield. For other traits, the medium production level has the highest cows number per common sires.For milk yield, the average age in the low production group was larger than in the other levels. Coefficient of determinations by production levels for each trait (using PROCFASTCLUS) were higher than 0.70.PROC FASTCLUS caused that within group’s variances decreased to minimum, and maximized differences between groups; therefore, coefficient of variance in each group was low (Table 1).
|
Table 1. Descriptive statistics of data sets for milk production traits in different production levels (PL) |
||||
|
|
PL |
|
||
|
Parameter |
Low |
Medium |
High |
Total |
|
Milk yield |
|
|
|
|
|
TD records, no. |
87431 |
261269 |
478598 |
827295 |
|
Means ±SD (kg) |
19.9 ±2.42 |
26.7 ±1.54 |
31.2 ±1.56 |
27.5 ±4.08 |
|
CV (%) |
12.1 |
5.76 |
5.00 |
14.8 |
|
Cows, no. |
10666 |
30854 |
56616 |
98136 |
|
Common sire, no. |
373 |
373 |
373 |
373 |
|
Cows. /common sire, no. |
28.6 |
82.7 |
151.8 |
263 |
|
HY, no. |
420 |
951 |
775 |
2146 |
|
HTD, no. |
4782 |
10214 |
8003 |
22999 |
|
Age at first calving (avg), mo. |
30.2 |
26.9 |
26.6 |
27.0 |
|
Fat yield |
|
|
|
|
|
TD records, no. |
146561 |
415377 |
265114 |
827052 |
|
Means ±SD (g) |
683±94.7 |
911±61.4 |
1126±90.3 |
929±173 |
|
CV (%) |
13.9 |
6.74 |
8.02 |
18.7 |
|
Cows, no. |
17916 |
53410 |
33807 |
105133 |
|
Common sire, no. |
366 |
366 |
366 |
366 |
|
Cows. /common sire, no. |
48.9 |
145 |
92.4 |
287 |
|
HY, no. |
444 |
1104 |
658 |
2206 |
|
HTD, no. |
4847 |
12025 |
7781 |
24653 |
|
Age at first calving (avg), mo. |
27.5 |
26.9 |
26.5 |
26.9 |
|
Protein yield |
|
|
|
|
|
TD records, no. |
46016 |
326381 |
217933 |
590330 |
|
Means ±SD (g) |
663±102 |
863±48.5 |
1023±71.3 |
891±135 |
|
CV (%) |
15.4 |
5.62 |
6.97 |
15.2 |
|
Cows, no. |
6845 |
42802 |
28325 |
77972 |
|
Common sire, no. |
324 |
324 |
324 |
324 |
|
Cows. /common sire, no. |
21.1 |
132 |
87.4 |
240 |
|
HY, no. |
263 |
950 |
589 |
1802 |
|
HTD, no. |
2199 |
9524 |
6637 |
18360 |
|
Age at first calving (avg), mo. |
27.9 |
26.8 |
26.6 |
26.8 |
|
Fat % |
|
|
|
|
|
TD records, no. |
199931 |
376226 |
136896 |
713053 |
|
Means ±SD (%) |
3.04 ±0.137 |
3.36 ±0.0831 |
3.67 ±0.131 |
3.41 ±0.248 |
|
CV (%) |
4.51 |
2.47 |
3.57 |
7.27 |
|
Cows, no. |
24328 |
46853 |
16747 |
87928 |
|
Common sire, no. |
405 |
405 |
405 |
405 |
|
Cows /common sire, no. |
60.1 |
115 |
41.4 |
217 |
|
HY, no. |
643 |
1237 |
515 |
2395 |
|
HTD, no. |
6225 |
12131 |
5066 |
23422 |
|
Age at first calving (avg), mo. |
26.8 |
27.0 |
27.0 |
26.9 |
|
Protein % |
|
|
|
|
|
TD records, no. |
165047 |
357267 |
130133 |
652446 |
|
Means ±SD (%) |
2.92 ±0.0950 |
3.11 ±0.0549 |
3.36 ±0.133 |
3.07 ±0.142 |
|
CV (%) |
3.25 |
1.77 |
3.96 |
4.63 |
|
Cows, no. |
21319 |
43911 |
16009 |
81239 |
|
Common sire, no. |
430 |
430 |
430 |
430 |
|
Cows. /common sire, no. |
49.6 |
102 |
37.2 |
188 |
|
HY, no. |
403 |
1162 |
151 |
1716 |
|
HTD, no. |
4173 |
12333 |
1648 |
18154 |
|
Age at first calving (avg), mo. |
26.3 |
26.8 |
27.0 |
26.2 |
Variance components and heritabilities
The sire additive genetic, permanent environmental and residual variances for 305-day production were heterogeneous among production levels, but the sire additive genetic variances of medium and high levels for protein percentage were similar and lower than sire additive variances in low production level (Table 2).
 |
Table
3 reflected (co)variance components for random regression coefficients for
calculating of sire additive genetic, permanent environmental and
residual variances (![]() respectively) of each trait in different PL. Residual variances for all
traits in the low production level were lower than inother levels.
Difference in variance component values between PL indicated that there were
differences in heritabilities between PL. Heritabilities for milk production
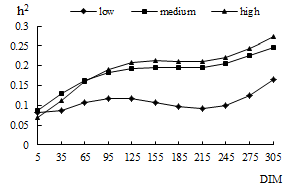
traits as a function of DIM in the three levels of production are shown in
Figure 1. Heritabilities for milk yield by DIM were higher than for other
traits. These heritabilities are in accordance with other studies (Shadparvar
and Yazdanshenas 2005; Abdullahpour et al 2010). Generally, heritabilities
for milk production traits were the highest in the high production level,
except for protein percentage. The highest heritabilities for protein
percentage in low PL was due to highest sire additive genetic variances.
respectively) of each trait in different PL. Residual variances for all
traits in the low production level were lower than inother levels.
Difference in variance component values between PL indicated that there were
differences in heritabilities between PL. Heritabilities for milk production
traits as a function of DIM in the three levels of production are shown in
Figure 1. Heritabilities for milk yield by DIM were higher than for other
traits. These heritabilities are in accordance with other studies (Shadparvar
and Yazdanshenas 2005; Abdullahpour et al 2010). Generally, heritabilities
for milk production traits were the highest in the high production level,
except for protein percentage. The highest heritabilities for protein
percentage in low PL was due to highest sire additive genetic variances.
|
Table 3. (Co)variance components for random regression coefficients of sire additive genetic (G), cow permanent environmental (PE) and residual (R) variances of each trait in different production levels (PL) with multiple-trait random regression sire model |
|||||||||||||||||||||
|
Parameters |
|
Milk yield (kg) |
|
Fat yield (g) |
|
Protein yield (g) |
|
Fat % (×10-3) |
|
Protein % (×10-3) |
|||||||||||
|
PL |
Low |
Medium |
High |
Low |
Medium |
High |
Low |
Medium |
High |
Low |
Medium |
High |
Low |
Medium |
High |
||||||
|
G |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
a0 |
a0 |
|
0.910 |
2.80 |
3.40 |
|
1232 |
1536 |
2412 |
|
1056 |
1975 |
2760 |
|
5.26 |
13.2 |
18.8 |
|
4.15 |
3.07 |
3.06 |
|
a0 |
a1 |
|
0.103 |
0.360 |
0.604 |
|
80.8 |
291 |
448 |
|
263 |
368 |
623 |
|
1.11 |
1.87 |
2.15 |
|
0.51 |
0.47 |
0.42 |
|
a0 |
a2 |
|
-0.0483 |
-0.139 |
-0.236 |
|
-27.5 |
28.5 |
50.2 |
|
-23.4 |
-104 |
-122 |
|
-0.05 |
-0.82 |
-1.13 |
|
-0.08 |
-0.12 |
-0.01 |
|
a0 |
a3 |
|
0.0949 |
0.123 |
0.224 |
|
13.6 |
8.87 |
-9.52 |
|
-16.3 |
34.3 |
122 |
|
-0.06 |
-0.07 |
0.47 |
|
0.03 |
-0.04 |
-0.07 |
|
a1 |
a1 |
|
0.122 |
0.187 |
0.292 |
|
109 |
174 |
235 |
|
510 |
180 |
322 |
|
0.60 |
1.01 |
1.30 |
|
0.73 |
0.37 |
0.55 |
|
a1 |
a2 |
|
-0.0258 |
-0.0322 |
-0.0509 |
|
1.65 |
2.46 |
23.3 |
|
-29.6 |
-28.7 |
-2.93 |
|
0.13 |
-0.35 |
-0.46 |
|
-0.11 |
-0.05 |
0.04 |
|
a1 |
a3 |
|
0.0158 |
0.0142 |
0.0459 |
|
-18.4 |
-3.38 |
-7.34 |
|
-26.6 |
4.25 |
23.3 |
|
-0.05 |
0.24 |
0.48 |
|
0.04 |
0.00 |
-0.04 |
|
a2 |
a2 |
|
0.0386 |
0.0326 |
0.0544 |
|
49.7 |
23.9 |
37.1 |
|
56.8 |
26.0 |
29.3 |
|
0.21 |
0.29 |
0.45 |
|
0.09 |
0.05 |
0.24 |
|
a2 |
a3 |
|
-0.0176 |
-0.0139 |
-0.0249 |
|
-15.9 |
-15.8 |
-7.02 |
|
-4.47 |
-2.32 |
-7.27 |
|
-0.09 |
-0.17 |
-0.33 |
|
-0.01 |
0.01 |
-0.02 |
|
a3 |
a3 |
|
0.0264 |
0.0225 |
0.0320 |
|
22.6 |
19.1 |
14.6 |
|
23.3 |
14.1 |
15.3 |
|
0.23 |
0.21 |
0.61 |
|
0.04 |
0.04 |
0.14 |
|
PE |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
p0 |
p0 |
|
19.1 |
33.6 |
38.0 |
|
22100 |
28830 |
39990 |
|
20000 |
26240 |
32970 |
|
99.9 |
134 |
152 |
|
36.9 |
28.4 |
29.0 |
|
p0 |
p1 |
|
0.949 |
1.73 |
2.86 |
|
-371 |
98.5 |
1154 |
|
2010 |
2701 |
3522 |
|
8.29 |
14.1 |
13.5 |
|
3.04 |
3.12 |
2.74 |
|
p0 |
p2 |
|
-1.09 |
-2.08 |
-2.08 |
|
-455 |
-621 |
-1124 |
|
-1827 |
-1423 |
-1577 |
|
-0.60 |
-1.43 |
-3.08 |
|
-0.05 |
0.65 |
-0.67 |
|
p0 |
p3 |
|
-0.216 |
0.119 |
0.540 |
|
-3.69 |
-103 |
-154 |
|
436 |
-88.4 |
23.3 |
|
-7.99 |
-5.07 |
-3.95 |
|
-1.51 |
-1.21 |
-0.46 |
|
p1 |
p1 |
|
3.23 |
5.30 |
6.17 |
|
3326 |
5527 |
6516 |
|
252 |
4598 |
5829 |
|
29.1 |
35.7 |
34.9 |
|
9.11 |
8.71 |
8.03 |
|
p1 |
p2 |
|
-0.326 |
-0.660 |
-0.661 |
|
-1661 |
-1626 |
-1412 |
|
-184 |
-367 |
-542 |
|
-7.64 |
-12.0 |
-11.5 |
|
-2.41 |
-1.88 |
-1.89 |
|
p1 |
p3 |
|
-0.206 |
-0.383 |
-0.295 |
|
421 |
-177 |
-378 |
|
40.7 |
-297 |
-314 |
|
8.65 |
7.89 |
12.0 |
|
2.31 |
2.93 |
3.55 |
|
p2 |
p2 |
|
1.483 |
2.06 |
2.34 |
|
1058 |
2344 |
2469 |
|
191 |
1855 |
2187 |
|
19.2 |
21.4 |
20.2 |
|
4.66 |
4.12 |
3.96 |
|
p2 |
p3 |
|
-0.249 |
-0.404 |
-0.538 |
|
-273 |
-1074 |
-1288 |
|
-40.3 |
-351 |
-522 |
|
-5.98 |
-9.77 |
-11.8 |
|
-2.30 |
-2.24 |
-1.17 |
|
p3 |
p3 |
|
0.797 |
1.10 |
1.23 |
|
208 |
1232 |
1349 |
|
30.4 |
822 |
1055 |
|
16.3 |
18.6 |
19.8 |
|
4.05 |
4.64 |
4.47 |
|
R |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
r1 |
r1 |
|
7.69 |
10.1 |
12.7 |
|
37040 |
39600 |
53410 |
|
13100 |
14620 |
17610 |
|
215 |
401 |
340 |
|
41.9 |
61.4 |
72.7 |
 |
 |
 |
| Figure 1a. Estimated heritability (h2) as a function of days in milk (DIM) in three levels of production for milk yield | Figure 1b. Estimated heritability (h2) as a function of days in milk (DIM) in three levels of production for fat yield | Figure 1c. Estimated heritability (h2) as a function of days in milk (DIM) in three levels of production for protein yield |
 |
 |
| Figure 1d. Estimated heritability (h2) as a function of days in milk (DIM) in three levels of production for fat percentage | Figure 1e. Estimated heritability (h2) as a function of days in milk (DIM) in three levels of production for protein percentage |