| Livestock Research for Rural Development 29 (6) 2017 | Guide for preparation of papers | LRRD Newsletter | Citation of this paper |
The study was designed to evaluate the immune response of chickens among different poultry management following challenge to live La Sota vaccine and further more investigated the lower antibody response detected in traditionally managed chicken, by testing the hypothesis that veterinary intervention (treatment with anti-helminthic, antibiotics and vitamin supplements), prior to vaccination, would improve the antibody response to live NDV vaccination.
Even if, a protective level of antibody (> 4 Log 2HAI) was detected in all chickens following vaccination, antibody titres were significantly higher (p < 0.05) in the intensively managed chickens Geometric Mean Titre (GMT) 85.4 when compared with the traditionally managed chickens GMT 59.3. From multivariable analysis; following a single live ND virus vaccine at the age of 21 day, chickens rearing in intensive production system were 8.6 times more likely to have high titre (128 vs 32) compared to those rearing in backyard free ranging system. The lower ND-HI titre in extensively managed chickens recorded in our study had be explained by impaired immune-competence due to immune-suppressive of concurrent infection. The study investigated the effect of concurrent parasitic and viral diseases on the immune response; as well as differences in protection among indigenous ecotype and exotic breed of chicken. The hypothesis that parasitism contributes to a poorer response to vaccination was proven (p < 0.05). Chickens that received veterinary treatment (de-wormed chickens) prior to vaccination were approximately 14 (i.e. 14.3) times more likely to have a titre of 128 when compared to titre 32 showing evidence for veterinary intervention had detectable effect on humoral response of chicken to vaccination (p< 0.05). Moreover; it was evident that HI titer differed between individual chickens and was influenced by breed types. Multivariate analysis showed that indigenous chickens were about 7 times more likely to have high HI titre (128 vs 32) compared to exotic breeds (p< 0.05). Though breeds differ genetically in their immune responsiveness to the antigen, and being protected following challenge; difference in their susceptibility to the virulent challenge virus needs to be ruled out in order to facilitate the improvement of disease-resistant traits.
Keywords: antibody titre, immune response
It is clear that poultry production is a growing and economically important industry and therefore the interest in improving the production results through improved health of the poultry is great. Though the epidemiology of ND is not clearly understood in village chickens of Ethiopia, field reports indicate that ND seems the most important disease epidemic inflicting heavy losses (Zeleke et al 2005). There are many recent reports of considerable economic loss due to mortality and cost of control of the disease (Leslie 2000; Capua et al 2002). ND is listed on Office International des Epizooties (OIE) list A of contagious diseases (OIE 2000) and furthermore, it is also a great threat to the third world where it is an endemic disease and one of the limiting factors for the poultry production (Awan et al 1994; Alexander 2001). Though the impact of ND in chickens reared on cage systems have diminished due to modernization in poultry farming, bio-security, culling of infected birds (Marangon and Busani 2006; Seal et al 2000) and appearance of virulent ND viruses (vNDV) in different parts of the world (OIE 2008) requires repeated and expensive use of live and/or inactivated vaccines (Van Eck 1987). The goal of vaccination is always sterilizing immunity, however, that has not yet been achieved with NDV vaccines. At best, NDV vaccines induce an immune response that reduces or completely prevents clinical disease and mortality from ND, decreases the amount of vNDV shed into the environment, and increases the amount of virus needed to infect the vaccinated animal (Marangon and Busani 2006 and Miller et al 2009). An interesting alternative to standard disease control methods would be selective breeding to increase disease resistance in livestock. However; chickens maintained on deep litter system and backyard free ranging system still remain susceptible to Newcastle disease via failure of effective immune response to ND vaccination. In addition; genetic resistance to disease involves many facets of the body’s defence system and interactions, and is extremely complex (IlayaBharathi et al 2016). Disease resistant traits had low heritability, which made it difficult to use conventional breeding to improve the disease resistant traits.
Lentogenic strains like La Sota are immunogenic enough and widely used all over the world, but still have variable residual pathogenicity (Borland and Allan 1980). Mesogenic strains are certainly more immunogenic (Al-Garib et al 2003) but used only in countries where ND is endemic. Both cause severe vaccine reactions that could be partly explained by their pathogenicity and the paucity of phagocytes in avian respiratory system and therefore lower activity against concurrent bacterial infections (Toth 2000).
Outbreaks of ND are unpredictable and discourage villagers from paying proper attention to the husbandry and welfare of their chickens (Alders et al 2012). The spread of ND virus within and between village poultry populations is relatively slow due to a low contact rate. The major mode of transmission of ND appears to be by the faecal-oral route. There is also report that some of the ecto-parasites, especially tick and mites, are vectors of other poultry diseases such as Newcastle disease, Fowl pox, and possibly chlamydia (Ruff 1999).
ND depends upon a number of factors such as NDV pathotype, age, structure and immunity of the host population, birds management setting either in intensive husbandry (such as in a commercial poultry farm) or in a traditional rural farm, breed susceptibility, concurrent disease and seasonal influences (Awan et al 1994).
It has been reported that parasitic infection or their concurrent infections result in immune suppression, especially in response to vaccines against some poultry diseases (Permin et al 2000). According to Glukhov`s (1984) study on the effect of A.galli infection on immunity to ND, the antibody level declines more rapidly in infected chickens.
The most important step in the process of improving the health and survival rate of the poultry is to get an extended and more detailed knowledge of how the immune system responds to the vaccine caused by the different pathogens.
New opportunities to improve our understanding of the genetic nature of disease resistance have been produced through recent advances in molecular biology, gene mapping, and immunology which make selection for disease resistance possible in the future. However, testing and selection for disease resistance and improved immune responsiveness require knowledge of the genetic correlations between disease resistance and immune responsiveness, and production traits. Some groups of Thai indigenous chicken have survived H5N1 outbreaks, and are considered to be resistant. These traits are related to immunogenetics, in particular.
The aim of this study was to examine the response to NDV vaccination in introduced birds managed either in intensive husbandry (such as in a commercial poultry farm) or in a traditional rural farm based management setting. This study confirmed that the village reared birds have a poorer response to vaccination. The hypothesis that concurrent infection contributes to a poorer response to vaccination was examined in the second study.
Studies were conducted within 8 km radius of the town DebreZeit in Central Ethiopa. DebreZeit is located at the geographical co-ordinates 8° 45' 0" N, 38° 59' 0" E, with an elevation of 1902m above sea level. Chickens were reared using one of two production systems, either intensive conditions on a commercial poultry farm or reared with limited management by small-holder farmers in a village setting.
The first study compared the antibody response of intensively and traditionally managed commercial chickens to live NDV vaccination.
Sixty-three, 21 day old chicks (Bovance brown breed) were sourced from a commercial poultry farm in Ethiopia. All chicks were vaccinated with NDV vaccine, (LaSotastrain), via eye drop, as outlined by the manufacturer (National Veterinary Institute/NVI, Ethiopia), at 21 days of age, receiving one drop intraocular applied using an eye-dropper calibrated according to the calibration methods described by Alders and Spradbrow (2001).
Vaccinated chicks were randomly assigned to one of two groups and managed under commercial or tradition management system for 15 days (Groups A and B, Table 1). To monitor the antibody response to vaccination, chickens were bled from the wing vein at 36 days of age, 15 days after vaccination.
The second study investigated the effect of prophylactic veterinary intervention (anti-parasitic, supportive treatment and antibiotics) on the immune response elicited to the live NDV vaccine in both indigenous and commercial chickens reared in a traditional scavenger management system.
Seventy chickens (commercial and indigenous breeds), reared under a traditionally, scavenger managed system were selected from 17 households in two villages. Chickens had not received any prior vaccinations or veterinary treatments. Chickens were randomly assigned into one of four treatment groups (Group C to F, Table 1).Chickens in groups D and F received veterinary intervention in the 15 days prior to vaccination with live NDV vaccination (LaSota strain). The anti-helmintic agents Piperazine (15 mg/ kg) and Amprolium 20% (0.3 g/l water) were administer orally daily for 13 days prior to vaccination and Levamisol (35 mg/kg) for 2 days prior to vaccination. Multi-vitamins (Amino Vet) and the broad spectrum antibiotic Oxytetracycline 20% (0.5 g/l water) were also orally administered daily for 14 days and 7days; respectively, prior to vaccination.
Two days after the veterinary intervention was terminated, all chickens in Groups C, D, E and F were vaccinated with live NDV vaccine as described previously. To monitor the antibody response to vaccination, chickens were bled from the wing vein at approximately 11 days after the single vaccination.
All chickens were seronegative to NDV, as determined by haemagglutination inhibition assay at the start of the studies/Day 0.
|
Table 1. Treatment groups |
||||
|
N |
Breed |
Husbandry |
Pre-vaccination |
|
|
Study 1 |
||||
|
Group A |
36 |
Commercial |
Commercial |
N/A |
|
Group B |
27 |
Commercial |
Traditional1 |
N/A |
|
Study 2 |
||||
|
Group C |
17 |
Commercial |
Traditional |
None |
|
Group D |
26 |
Commercial |
Traditional |
Yes |
|
Group E |
16 |
Indigenous |
Traditional |
None |
|
Group F |
11 |
Indigenous |
Traditional |
Yes |
Antibodies against NDV were determined by using the haemagglutination inhibition (HAI) assay (Beard 1989 and OIE 2000). Briefly, two-fold serial dilutions of serum samples dilution in phosphate buffered saline were incubated for 45 mins at room temperature with 4 Haemagglutination Units of NDV (freeze-dried vaccine with a minimum titer of 10p7 ELD50. (ELD = embryonic lethal dose)), followed by the addition of 1% chicken red blood cells. The HAI titre of each sample was expressed as the reciprocal of the highest dilution of serum that completely inhibited Haemagglutination of the red blood cells, with HAI titres greater than 8 considered positive and titres of at least 32 considered protective (Allan and Gough 1974).
Treatment groups and data for sero-protection were analysed by Logistic regression using SPSS. For all quantitative data a P-value <0.05 was considered as statistically significant.
These studies were conducted to evaluate the immune response of chickens among different poultry managements following challenge to LaSota vaccine. Most ND vaccines are live vaccines. In the current study, a protective level of antibody (> 4 Log2 HAI) was detected in all experimental chickens following a single vaccination, irrespective of management type. Antibody titres were, however, significantly higher (p < 0.05) in the intensively managed chickens GMT 85.4 when compared with the traditionally managed chickens GMT 59.3. Field results suggest that only birds with HI titers greater than > 4 Log2 HAI after multiple vaccinations will survive a vNDV challenge as 66% of the flock succumbed with titers less than that (Kapczynski and King 2005). More commonly, HI levels of 32 or higher (≥4 Log2 HAI) are typically thought of being protective (Allan et al 1978). Although there are no published reports about the differences of NDV antibodies among different managements in Ethiopia; based on the laboratory investigations and prospective follow up of experiment chickens, the authors were convinced that the immune response of exotic chickens against ND vaccine would be reduced whenever the chickens are reared in backyard production system. From logistic regression analysis, the study showed that following a single live NDV vaccine at day 21, chickens rearing in intensive production system were 8.6 times more likely to have high titre (128 vs 32) compared to those rearing in backyard free ranging system (Table 3). In non-immune, intensively managed chickens; vaccination of ND vaccine in appropriate manufacturer dose is in itself a sufficient level to induce antibody production in order to provide sero-protection against vNDV if the vaccines are viable, administered correctly to healthy birds and time is allowed for an appropriate immune response to develop prior to exposure to the challenge virus (Cornax et al 2012, Dortmans et al 2012 and Kapczynski and King 2005). In traditionally-managed chickens, however, vaccination may not induce sufficient amount of antibody production. Despite the nature and epidemiology of immune response against NDV in backyard free ranging system poorly understood, the lower ND-HI titre in extensively managed chickens recorded in our study had be explained by impaired immune-competence due to immune-suppressive of concurrent infection or by immune-suppressive substances in the feed. Moreover; availability of sufficient feed, stress, favourable physical environment and heterogeneity of NDV probably involved in immune-suppression and thereby influences immune response variation among different poultry management systems. The hypothesis that concurrent infection contributes to a poorer response to vaccination was confirmed in the subsequent study as reported below (Table 4). Many studies reported that various nutrients have a significant influence on the immune system of poultry. For example, the shortage of valine (it makes 7% of poultry gammaglobuline) reduces the HI titre (Bhargava et al 1970). According to Mazija et al (1981), the total amount of proteins in feed has significant effects on the immune response to ND. The nutritional management practiced in the traditionally managed chickens was predominantly scavenging which is unbalanced diet and responsible for failure of immune response.
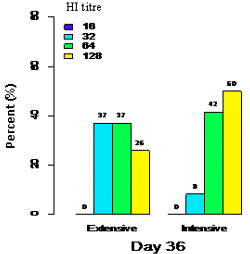
Haemagglutination inhibition antibody titres and GMT for intensively- and traditionally managed commercial chickens following live NDV vaccination are presented in Figure 1 and Table 2, respectively.
|
|
| Figure 1. Antibody titres among different management following vaccination with La Sota vaccine on days 21 and bleeding on day 36. |
|
Table 2. Geometric mean antibody titres (GMT) among chickens managed under different management setting following vaccination with live LaSota vaccine. |
|||
|
Management system |
No of chickens |
GM HAI antibody titres |
Level of protection |
|
Intensive |
36 |
85.4 |
Protective |
|
Traditional |
27 |
59.3 |
Protective |
|
Table 3. Multivariable analysis of HI titer with poultry production system |
|||||
|
HI at Day 36 |
Coeff/B |
SE |
P |
OR |
|
|
128 v’s 32 |
Intensive |
2.15 |
0.79 |
0.007 |
8.6 |
This experiment was designed to investigate the lower antibody response detected in traditionally managed chicken, by testing the hypothesis that veterinary intervention (treatment with anti-helmintics, antibiotics and vitamin supplements), prior to vaccination, would improve the antibody response to live NDV vaccination. An analysis of the HI antibody titres detected in the different treatment groups is outlined in Table 4.
In this study, a sero-protective HI antibodies were detected for all but one chicken (Group C, HI antibody titre 16), 15 days following live NDV vaccination (figure 2). Irrespective of breed, the treated groups (Groups D, HAI GMT 112 and Group F, HAI GMT 114) tended to have higher antibody levels than the controlled/untreated (Groups C HAI GMT 50 and Group E HAI GMT 76).
 |
 |
| Figure 2. Antibody titre in different breed and treatment groups of experiment animals | |
Chickens that received veterinary treatment prior to vaccination were approximately 14 (i.e. 14.3) times more likely to have a titre of 128 when compared to titre 32. Hence, the veterinary intervention described in this study increased the probability that the chicken would have a stronger humoral response to vaccination (Table 4).
In general significant association was found between the parasite burden, breed/ecotype, and the HI-titer measured during the experimental period. This was consistent with previous studies that demonstrate a significant association of high helminthes burden and low antibody response (Sofus 2003). Concurrent infection in free range chickens is very common. Lack of efficiency of vaccines against ND has been reported (Spradbrow 1990) and previous investigations have indicated that infection with endoparasites can reduce the immune response towards ND vaccines (Bhopale et al 1998, Hørning et al in press). According to Perozo et al (2012) the presence of immunosuppressive organisms can also render ineffective a protocol that is sound under experimental conditions. Indeed even the most efficacious vaccine cannot induce an immune response if the bird is immunosuppressed. Helminths and viral infections (ND vaccine) are known to induce a Th2 immune response in murine. At the same time helminths contribute to an immunosuppressive effect on the immune response against viral infections in chickens. Literatures showed that individuals with concurrent helminths had significantly lower number of lymphocytes (Dahl et al 2002), which in turn suppresses the immune response against viral infection in which the lymphocytes are a key factor (Agger et al 1995).
|
Table 4. Multivariate analysis of HI antibody titer with breed and parasitic burden |
||||
|
Titre
|
B |
SE |
p |
OR |
|
Intercept |
-3.53 |
1.35 |
0.009 |
|
|
Breed |
1.94 |
0.87 |
0.03 |
7.0 |
|
Treatment |
2.66 |
0.82 |
0.001 |
14.3 |
Despite limited sample size for the current experiment, from multivariate logistic regression analysis, it was evident that the level of antibody (HI titer, 128 vs 32) differed between individual chickens and was influenced by breed (i.e. exotic or indigenous) (Table 4). Specifically, indigenous chickens were about 7 times more likely to have high HI titre (128 vs 32) compared to exotic breeds. Breed comparative to ND antibody response profile, have not yet been isolated in chickens, and only few papers deal with concurrent infections in chickens. However, some studies tried to highlight indigenous chicken were known to possess desirable characteristics such as thermo-tolerance, resistance to some diseases, despite their low productivity. It is clear that the breed difference had a pronounced effect on weight gain, white blood cell differential counts. Even if, the hypotheses may not have been the underlying explanation requires further investigation, weight gain and white blood cell differential counts may associate with the disease resistance difference. Similarly, according to previous work conducted by Hassan et al (2004) and King (1996), genetic resistance to ND has been observed with various lines among breeds of chickens. Concerning this topic it is important to note that each Newcastle disease virus may be better adapted to grow in one species versus another. In addition, it is known that there is a negative correlation between a primary antibody response to NDV and favorable production traits (Lwelamira et al., 2009).
The authors are greatly indebted to the CH4D project for attachment of my MSc thesis research work and providing technical support during the courses of this study. In addition; authors would like to thank farm households that collaborate during sample collection for the study.
Agger R, Leslie G and Aasted B 1995 Immunology textbook.2nd ed. pp. 1-292.
Alders R, Spradbrow P B 2001 Controlling of Newcastle disease in village chickens. A field manual, Australian Centre for International Agriculture Research (ACIAR), Research monograph No. 82: 1 – 112.
Alders R, Bagnol B, Costa R and Young M P 2012 Sustainable Control of Newcastle Disease in Village Poultry. INFPD Good Practices for Family Poultry Production Note No 05.
Alexander D J 2001 Newcastle disease (APMV-1). In: F. Jordan, M. Pattison, D.J. Alexander and T. Faragher (eds), Poultry Disease, 5th edn, (Harcourt Publishers, London), 259-268
Al- Garib S O, Gielkens A L J, Gruys E and Koch G 2003 Review of Newcastle disease virus with particular references to immunity and vaccination. Worlds Poult Sci J 59: 185-200
Allan W H and Gough R F 1974 A standard haemagglutination inhibition test for Newcastle disease: a macro and micro methods. Vet. Rec., 95: 120-123
Allan W H, Lancaster J E and Toth B 1978 Newcastle disease vaccines-Their production and use. FAO Animal Production and Health Series No. 10. FAO: Rome, Italy.
Awan M A, Otte M J and James A D 1994 The epidemiology of Newcastle disease in rural poultry: a review. Avian Pathology, 23, 405–425
Beard C W 1989 Serologic procedures, p. 192-200. In H G. Purchase, L H Arp, C H Domermuth and J E Pearson (ed.), A laboratory manual for the isolation and identification of avian pathogens, 3rd ed. Kendall/Hunt Publishing Company, Dubuque, Iowa.
Bhargava K K, Hanson R P and Sunde M L 1970 Effects of methionine and valine on antibody production in chicks infected with Newcastle disease Virus. Journal of Nutrition vol. 100 pp 241-248.
Bhopale S T, Deore M D, Gatne M L and Narsapur V S 1998 Immunosuppression in birds experimentally infected with single and mixed parasitic infections. Journal of Bombay veterinary College vol. 6 pp 17-19
Borland L J and Allan W H, 1980 Laboratory tests for comparing live lentogenic Newcastle disease vaccines. Avian Pathology vol. 9 pp 45-59
Capua I, Dalla P M, Mutinelli F, Marangon S and Terregino C 2002 Newcastle disease outbreaks in Italy during 2000: Vet Rec 150: 565-568
Cornax I, Miller P J and Afonso C L 2012 Characterization of live LaSota vaccine strain-induced protection in chickens upon early challenge with a virulent Newcastle disease virus of heterologous genotype. Avian Dis., Volume 56, pp. 464–470
Dahl C, Permin A, Christensen J P, Bisgaard M, Muhairwa A P, Petersen K M, Poulsen J S and Jensen A L 2002 The effect of concurrent infections with Pasteurella multocida and Ascaridia galli on free range chickens. Vet.Microbiol. 86(4): 313-324.
Dortmans J C, Peeters B P and Koch G 2012 Newcastle disease virus outbreaks: vaccine mismatch or inadequate application? Vet. Microbiol., Volume 160, pp. 17–22
Glukhov E P 1984 The effect of ascaridiasis on post-vaccination immunity of chickens to ND, parazitotsenozyIassotsiztivnye, Bolezni, 139-142
Hassan M E, Smith G W, Ott R S, Faulkner D B, Firkins L D, Ehrhart E J and Schaeffer D J 2004 Reversibility of the reproductive toxicity of gossypol in peripubertal bulls. Theriogenology, 61 (6): 1171-1179
Hørning G, Rasmussen S, Permin A and Bisgaard M In Press: Investigations on the influence of helminth parasites on vaccination against Newcastle disease virus under village conditions
IlayaBharathi D, Raj Manohar G and Shamsudeen P 2016 Selection methods for disease resistance in poultry: a review. International Journal of Science, Environment, and Technology, Vol. 5, No 2, 810-815
Kapczynski D R and King D J 2005 Protection of chickens against overt clinical disease and determination of viral shedding following vaccination with commercially available Newcastle disease virus vaccines upon challenge with highly virulent virus from the California 2002 exotic Newcastle disease outbreak Vaccine, Volume 23, 2005, pp. 3424-3433
King D J 1996 Influence of chicken breed on pathogenicity evaluation of velogenic neurotropic Newcastle disease virus isolates from cormorants and turkeys Avian Dis., Volume 40, 1996, pp. 210–217
Leslie J 2000 Newcastle disease: outbreak losses and control policy costs. Vet Rec 146: 603-606
Lwelamira J, Kifaro G C and Gwakisa P S 2009 Genetic parameters for body weights, egg traits and antibody response against Newcastle disease virus (NDV) vaccine among two Tanzania chicken ecotypes Trop. Anim. Health Prod., Volume 41, 2009, pp. 51–59
Mazija H, Bidin Z, Štajer A and Mikec M 1981 Vakcinacija iz nužde kokoši protiv atipične kuge peradi (Emergency vaccination against Newcastle disease). Praxis vet 29: 45-52
Marangon S and Busani L 2006 The use of vaccination in poultry production. Rev. Sci. Tech., Off. Int. Epiz., 26(1): 265-274.
Miller P J, Estevez C, Yu Q, Suarez D L and King D J 2009 Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis., Volume 53, pp. 39–49
Office International des Epizootics 2000 Newcastle Disease. In: Manual of Standard for Diagnostic Tests and Vaccines.5th edition, Paris, Pp. 104-124.
OIE 2008 World animal health information disease (WAHID) interface. (www.oie.int/wahid-prod/public. php?page=weekly_report_index&admin=0)
Permin A, Pedersen G and Riise J C 2000 Poultry as a Tool for Poverty Alleviation: Opportunities and Problems Related to Poultry Production at Village Level. SADC planning workshop on Newcastle disease control in village chickens, Maputo, Australian Centre for International Agriculture Research, ACIAR proceedings No. 103: 143-147
Perozo F, Marcano R and Afonso C L 2012 Biological and phylogenetic characterization of a genotype VII Newcastle disease virus from Venezuela: efficacy of field vaccination J. Clin. Microbiol., Volume 50, 2012, pp. 1204-1208
Ruff M D 1999 Nematodes and Acanthocephalans. In: Hofstad M.S., Barnes H.J., Calnek .W., Reid W.M. &Yorder H.W.(eds.): Diseases of poultry 8th ed. Iowa State University Press, Ames. pp. 614-648.
Spradbrow P B 1990 Village poultry and preventive veterinary medicine. Preventive Veterinary Medicine, 8, 305– 307
Seal B S, King D J, Sellers H S 2000 The avian response to Newcastle disease virus, Dev. comp. immunol.,Volume 24, 2000, pp. 165-170
Sofus B 2003 Avian immune response in relation to Newcastle disease in parasite infected chickens pp. 1-160
Tadesse S 2000 Prevalence of Avian Tuberculosis in Three Selected Agro-climatic Zones of Central Ethiopia. [Veterinary doctoral thesis] Faculty of Veterinary Medicine Addis Ababa University DebreZeit Ethiopia. Pp. 21-29
Toth T E 2000 Nonspecific cellular defence of the avian respiratory system: a review. Dev Comp Immunol 24:121-139
Van Eck J H 1987 Immunity to Newcastle disease in fowl of different breeds, primarily vaccinated with commercial inactivated oil-emulsion vaccines: a laboratory experiment. 9: 296-303
Zeleke A, Sori T, Gelaye1 E and Ayelet G 2005 Newcastle Disease in Village Chickens in the Southern and Rift Valley Districts in Ethiopia. International Journal of Poultry Science. 4 (7): 507-510
Received 11 January 2017; Accepted 14 March 2017; Published 1 June 2017