a,b,c values in each row with different superscripts are significantly different (p < 0.05) by using ANOVA post hoc (Tukey test).
| Livestock Research for Rural Development 29 (4) 2017 | Guide for preparation of papers | LRRD Newsletter | Citation of this paper |
This study assessed the effects of hyperthermal –induced stress on survival and expression of heat shock protein in Nile tilapia,Oreochromis niloticus fingerlings fed aqueous extract from Moringa oleifera leaf. Fish were graded by size and groups of 15 fish of 6.00 ± 0.10g and then stocked into glass tanks of 60cm ×45cm×45cm dimension in triplicate. They were fed diets representing different supplementation levels of Moringa oleifera leaf extract. The graded levels of M. oleifera leaf extract were 0.00g (control), 0.05g, 0.10g, 0.15g, 0.20g, 0.25g per 100g for each diet denoted as MLST0, MLST5, MLST10, MLST15, MLST20 and MLST25. Six isonitrogenous and isocaloric diets were formulated to meet the requirements of 30% crude protein. After six weeks of the feeding trial, fish previously fed each experimental diet were exposed to hyperthermia – induced stress. Water temperature was maintained at 37 ± 0.500C throughout the hyperthermia – induced stress period. After hyperthermia – induced stress, mortality was monitored and molecular expression (abundance and diversity) of stress protein genes (heat shock protein, HSP70) in experimental fish was assessed.
Hyperthermia – induced stress mortality was reduced by supplementing the fish diets with M. oleifera leaf extract at 0.10g, 0.15g, 0.20g, 0.25g per 100g dosages. Molecular analyses of the HSP 70 genes showed marked similarity in the banding patterns of fish fed the Moringa leaf supplemented diets with a corresponding increase in fish survival in these treatments. Based on the result of this study, a dose of 0.10g/100g dietary Moringa leaf supplementation was sufficient as a stress reducing agent and improved the survival of fish.
Keywords: extracts, heat shock proteins expression, hyperthermia, mortality
Stressors in aquaculture are unavoidable and cause many harmful effects. Stress and stress-related diseases are currently a much discussed topic in animal including fish husbandry and research (Guo 2016). Cultured fish are unavoidably exposed to wide ranges of stimuli associated with environmental stress and pathological challenges (Xie et al 2008). All environmental factors can influence the degree to which fish respond to stressors. External factors include acclimation temperature, salinity, time of day, wave length of light and even background color of the tanks (Ojima et al 2005). Internal environmental factors, including the fish’s nutritional state and presence of disease may also affect the magnitude of the stress response (Blancheton et al 2013). Stress management is therefore becoming subjects of growing interest for an increasing number of aquaculture fish species (Conte 2004). Pathological, transportation and temperature stress, particularly rapid changes in temperature, severely limits the fish's ability to release antibodies, giving the invader the time to reproduce and overwhelm the fish (Giri et al 2015). Strategies to attenuate stress and stress-related negative responses in aquaculture should be considered. Using current knowledge in fish nutrition, the methods and criteria for optimal nutrient recommendations should be re-evaluated to include physiological responses and health factors such as immunology and disease resistance in fish (Wedemeyer et al 2000). There is also increasing concern about the effect of high temperature on aquaculture species due to climate change and aquaculture related heat stress (Ojima et al 2005). Knowledge and understanding of what constitutes stress in fish has increased immensely in the past few decades, notably in the area of physiological mechanisms and responses that lead to changes in metabolism and growth, immune functions, reproductive capacity and normal behavior (Li et al 2008). Moreover, understanding trends in changes that occur in fish in response to stressors can often provide clues that help relate the physiological responses of individuals with changes in performance that could affect their health and survivorship (Sen et al 2014). Primary stress responses in teleost fishes include a number of hormonal changes, particularly those in circulating levels of cortisol and catecholamines. Secondary responses, which may or may not be caused directly by the endocrine response, include measurable changes in blood glucose, lactate or lactic acid, and major ions (e.g., chloride, sodium, and potassium), and tissue levels of glycogen and recently HSPs (Heat Shock Proteins) (Liu et al 2015). Tertiary responses, includes changes in growth, disease resistance and behavior.
Different parts of Moringa have shown great antioxidant activity (Anwar et al 2007) as well as immunomodulatory function in animals (Ojiako 2014). Moringa leaves have been reported as a rich source of proteins, antioxidants and minerals. Extracts from the leaves and pods have numerous health benefits and seed extracts have been shown to be antimicrobial (Ojiako 2014). The leaves are abundant in the tropical areas and many studies have been conducted on their use but limited work has been done on their effects on aquaculture – related stress in Nile tilapia. Tilapia is considered as an excellent species for aquaculture in tropical and subtropical regions, as a result of the high tolerance to handling, stress situations and critical conditions, tolerance to sub optimal water quality, tolerance to high stocking densities, and its fast growth (Lim and Webster 2006). More than 22 tilapia species are cultured worldwide (FAO 2014). Therefore, due to the multiple role of Moringa oleifera Lam. (Moringaceae) in aquaculture (Rapatsa and Moyo 2014), a better understanding of the mechanism through which dietary M. oleifera as a nutritional supplement influences the physiology of fish exposed to stress in aquaculture especially at molecular and systemic levels is necessary. There are abundant information on how aquatic animals respond physiologically to general stress however little information is available on the understanding of how aquatic animals respond to thermal stress at the molecular level (Wang et al 2016). In order to provide a sustainable and environmental friendly solution to the adverse effects of stressors in Nile tilapia aquaculture, this study was designed to assess the effects of hyperthermia –induced stress on survival and expression of heat shock proteins in Nile tilapia fingerlings fed aqueous extract from Moringa oleifera leaf.
The leaves of Moringa oleifera were collected from a farm settlement at Ijare, Ondo State, Nigeria. It was identified and authenticated at the Department of Crop, Soil and Pest Management, Federal University of Technology, Akure. The leaves were destalked, washed and dried in the shade. Moringa oleifera leaves were ground with pestle and mortar, leaves were then extracted according to the modified method of Makanjuola et al (2013) as follows. Five hundred grams of the powdered leaf were soaked in 1.5 liter of warm water (60oC). Each solution was allowed to stand for 24 hours, after which it was sieved with a muslin cloth and filtered using No 1 Whatman filter paper. The filtrate were collected in a beaker and concentrated with the aid of rotary evaporator (Resona, Germany).
The feed ingredients were purchased at Adedom Feedmill, Ondo road, Akure, Ondo State, Nigeria. Six isonitrogenous and isocaloric diets were formulated to meet the requirements of 30% crude protein (Table 1) for O. niloticus fingerlings (National Research Council, 2010) using feed formulation software (WinFeed soft 2.0, USA). All dietary ingredients were weighed with a weighing top balance (Metler Toledo, PB8001 London). The ingredients were then ground to a small particle size (approximately 20 µg). Ingredients including Moringa oleifera extract, vitamin and mineral premix were thoroughly mixed in a Hobbart A-200T mixing machine (Hobbart Ltd London England) to obtain a homogenous mass. Alginate, Laminaria digitata (IGV GmbH, Germany®) was added as binder. The resultant mash was pressed without steam through a mincer using 2mm diameter die attached to the Hobbart pelleting machine. Diets were immediately air - dried, after drying the diets were broken up, sieved and stored in air-tight transparent plastic containers, labeled and stored until feeding. Standard and official methods (AOAC 1995) were used to perform the proximate analyses of feed of fish in the study.
|
Table 1. Composition of the experimental diet (g/100g) containing dietary Moringa Oleifera for Nile tilapia, Oreochromis niloticus fingerlings |
||||||
|
TILAPIA |
MLST0 |
MLST5 |
MLST10 |
MLST15 |
MLST20 |
|
|
Fish meal |
11.50 |
11.50 |
11.50 |
11.50 |
11.50 |
11.50 |
|
Groundnut cake |
19.50 |
19.50 |
19.50 |
19.50 |
19.50 |
19.50 |
|
Soybean meal |
20.50 |
20.50 |
20.50 |
20.50 |
20.50 |
20.50 |
|
Yellow maize |
24.50 |
24.50 |
24.50 |
24.50 |
24.50 |
24.50 |
|
Vegetable oil |
7.00 |
7.00 |
7.00 |
7.00 |
7.00 |
7.00 |
|
Rice Bran |
13.00 |
13.00 |
13.00 |
13.00 |
13.00 |
13.00 |
|
Alginate (Binder) |
2.00 |
1.95 |
1.90 |
1.85 |
1.80 |
1.75 |
|
Vitamin Mineral mix |
2.00 |
2.00 |
2.00 |
2.00 |
2.00 |
2.00 |
|
Moringa extract |
0.00 |
0.05 |
0.10 |
0.15 |
0.20 |
0.25 |
|
Composition of vitamin-mineral mix (Aquamix) (quantity/kg), Vitamin A, 55,00,000 IU; Vitamin D3, 11,00,000 IU; Vitamin B2, 2,000 mg; Vitamin E, 750 mg; Vitamin K, 1,000 mg; Vitamin B6, 1,000 mg; Vitamin B12, 6 mcg; Calcium; Pantothenate, 2,500 mg; Nicotinamide, 10 g; Choline Chloride, 150 g; Mn, 27,000 mg; I, 1,000 mg; Fe, 7,500 mg; Zn, 5,000 mg; Cu, 2,000 mg; Co, 450. L- lysine, 10 g; Selenium, 50 ppm. |
||||||
O. niloticus fingerlings were obtained from the Hatchery unit of the Department of Fisheries and Aquaculture Hatchery, Federal University of Technology Akure, prior to the feeding trial. Fish were graded by size and groups of 15 fish of 6.00 g per replicate for O. niloticus were stocked into glass tanks of 60cm ×45cm×45cm dimension. A commercial diet, Nutreco ® (35% crude protein) was fed to all fish during a 2- week conditioning period. Each experimental diet was fed to six replicate groups of fish for 70days. All groups were fed their respective diets at the same fixed rate (initially 5% of body weight per day). This rate was adjusted each week. Fish were fed by 0900-1000 and 1700-1800h GMT, for 7 days each week. Growth was monitored weekly by batch weighing of fish from each tank.
Physico-chemical water parameters : Dissolved oxygen was monitored using HANNA 98103SE (HANNA instruments, Rhode Island). Temperature and pH were monitored using YSI-IODO 700 Digital probe (IFI Olsztyn, Poland).
At the end of the feeding trial, 18 fish previously fed each experimental diet from each treatment representing 3 fish per treatment were kept in plastic tanks for hyperthermia- induced stress according to a modified method of Sen et al (2014) using a 2-kW heating rod (Binatone, Japan). The temperature treatments was based on a pilot study where the sub-lethal temperature of < 36oC was recorded for O. niloticus. The rate of heating ramp was about 3o C/h. Water temperature was maintained at 37 ± 0.5 0C throughout the hyperthermia – induced stress period. No fish died during the hyperthermia treatment. Two fish were taken randomly at 2h after exposure from the tanks. Two fish per tank were euthanized by overdose (200 mg / liter of water for 10 min) of tricaine methane sulphonate (MS222; Pharmaq, Fordingbridge, UK). Liver samples were removed by dissection and weighed immediately after hyperthermia- induced stress from fish for further analyses.
Extraction of DNA and PCR-TGGE of fish liver were performed using the Qiagen® blood + tissue through the spin column protocol, the gram positive optimisation was used in this kit to enhance the lysis of cells with complex cell walls. The liver was excised from the fish, homogenized and stored at -80oc to assess differences between the dietary groups. Extracted DNA was checked for quality and quantity by agarose electrophoresis and UV spectrophotometry using Nanophotometer P-class (IMPLEN®, USA).
Primers were sourced from literature and emphasis was laid on primers suitable for Temperature gradient gel electrophoresis (TGGE) (Table 2). PCR of the V3–5 region of the liver 16S rDNA genes was performed using primer 968-f (Ojima et al., 2005) as shown in Table 3. All were obtained from the same reputable supplier (Invitrogen, Germany). For extracted DNA, PCR was performed with a 15-mL reaction mixture containing 7.3mL Mango mix (Qiagen®), 1.5mM MgCl2 , 0.15 mL bovine serum albumin (BSA) which prevents reagents sticking to the tube, 0.75mL forward primer, 0.45mL reverse primer and 1.5mL of template DNA. Pure culture PCR was performed using Pyrogen free DNAse water of 4.9mL. Thermocycling conditions were as follows: initial denaturation at 94 oC for 2minutes 30 seconds, followed by 60oC for 30seconds at -0.7 per cycle for the annealing and elongation was at 72oC for 1min. Denaturation was done again at 94oC for 30 seconds for 15 cycles, then 49oC for 30 seconds and elongation at 72oC for 30 seconds. Final denaturation was done for 18 cycles and final elongation for 5 minutes and cooled down to 4oC.
Control gels and Quick load® DNA ladders (Biolabs, New England) were run on every gel. The condition for TGGE machine was at a constant voltage of 65 V for 16 h , the ramp temperature and increasing rate ramp (output change) was at 0.9oC per hour from 55.6 to 69.6 oC using a DCode system (BioRad). Photograph was documented with Gel documentation equipment, Gene genius by Syn gene® and processing was done using gene genius snap 6.0 software.
|
Table 2. Sequence and conditions of primers used in this study. PCR of the V3–5 region of the Nile tilapia targeting the 16S rDNA genes was performed using universal primers (Ojima et al 2005) for Nile tilapia. |
||||||||
|
No |
Author |
Oligo Name |
F primer name |
R primer name |
Sequence (5’-3’) |
Amplicon size |
Region |
GC-contents |
|
1 |
Ojima et al 2005 |
HP7A |
968-f |
140-r |
HSP70-F GGCAATGAGAGGTTCAGGTG HSP70-R TCTCATGGATGCCGCAGGA |
300 approx. |
V3 |
57.1%F 58.8%R |
This experiment was designed with a completely randomised design (CRD) to test for significant differences in the mean of treatments. The differences between mean of treatments were considered significant at P ≤ 0.05 by one way analysis of variance (ANOVA) using Statistica ® software. Follow–up procedures were performed where significant differences occurred in the means using Tukey test. Photographed gel after TGGE was processed for gel diversity and migration pattern using GelAnalyzer 2010a®, gel electrophoresis image analysis software. Minitab 18® statistical software was used to plot the web – profile radar plot for the diversity of the TGGE gels.
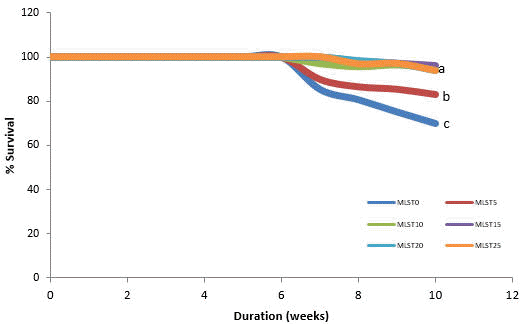
Significantly higher survival (ANOVA; F5, 12 = 483.01, P= 0.001) was found in fish fed MLST10 and MLST15 diets as 95.70 and 95.70 %, respectively at the end of the feeding trial. Reduction in % survival was recorded starting from week 7 which also correspond to the beginning of the stress challenge (Figure 1).
|
| Figure 1. Cumulative survival of O. niloticus fed the experimental diets for 10 weeks. a,b,c values in each row with different superscripts are significantly different (p < 0.05) by using ANOVA post hoc (Tukey test). |
Molecular analyses with the aid of TGGE gel showed similar banding patterns in fish fed the Moringa leaf supplemented groups in MLST10, MLST15, and MLST20 treatments (Figure 2). Each band represents one gene of HSP 70, the ladders are showed on the first lane of the TGGE gel. Dietary effects were observed in the banding patterns, with M .Oleifera based diets showing more bands on the banding profile on the gel compared with the control.
 |
| Figure 2.
PCR-TGGE gels showing diversity of the HSPs from the PCR-TGGE
fingerprints gel within the O. niloticus liver after the
feeding trials. Each lane represents sample from fish after subjection to A. hydrophila and hyperthermal –induced stress. Lane 1 = Ladder (arbitrary markers). |
A web-filled radar graph representing the similarity and richness of the HSPs from the PCR-TGGE fingerprints gel within the O. niloticus liver of the control and Moringa leaf supplemented groups at the end of the trial is presented in Figure 3. There were differences in the distribution of the banding profile of the HSP 70 in the liver of fish used in this study. The fingerprints gel showed that the MLST10, MLST15, MLST20 treatments were the most closely related which clustered into a group distinctly different to the profiles of the control, MLST5 and MLST25 groups.
 |
| Figure 3.
Web radar analysis with markers showing the distributions of the heat
shock protein genes of Oreochromis niloticus fed the experimental diets with intensity and molecular mass (kDa). |
Supplementation of Moringa leaf extracts increased the percentage survival of fish fed the experimental diets in the present study O. niloticus fed the control diet in this study had higher percentage mortality than fish fed the Moringa supplemented diets. This result is in agreement with the work of Tekle and Sahu (2015), who reported that the ameliorative effects of Moringa flower on O. niloticus subjected to Aeromonas hydrophila - induced stress was as a result of the ability of the plant extract in stimulating the activity of metabolic and antioxidative stress enzymes. M. oleifera plant has been widely reported to contain constituents such as phenolic acids, glycosides, kaempferol and quercetin (Ojiako 2014) which are believed to be responsible for enhancing immunity against oxidative stress and microbial diseases. In the present study, fish in MLST10 and MLST15 groups exhibited the highest post-challenge survival (95.70 %) and (95.13%) respectively. In agreement with the result of the present study, Wang et al (2015) reported that dietary administration of M. oleifera enhanced the post-challenge survival of common carp, Cyprinus carpio against A. hydrophila infection. Similarly, dietary administration of guava leaf at 30 mg/ kg for 45 days significantly enhanced the resistance of L. rohita to A. hydrophila infection (Giri et al 2015). The low percentage mortality in fish of MLST10 and MLST15 group might be associated with the improved resistance of fish against hyperthermia-induced stress resulting in higher post-challenge survival rates. The mechanism by which Moringa inhibits oxidation and mortality can be explained by the B ring hydroxyl configuration which is the most significant determinant of scavenging of ROS, because it donates hydrogen and an electron to hydroxyl, peroxyl, and peroxynitrite radicals, stabilizing them and giving rise to a relatively stable flavonoids radical (Kumar et al 2013). Hammed et al(2015) also reported that the presence of potent antioxidants in Moringa leaf can be correlated with increase in antibody production which helps in the survival and recovery of fish.
In the current study, supplementation with aqueous extract of Moringa leaves resulted in increased banding profile on the gel compared with the control suggesting that the inclusion of Moringa increase the accumulation of the HSPs in O. niloticus after A. hydrophila and hyperthermia –induced stress in the current study. In line with this result, Labeo rohita diet supplemented with aqueous extract of Psidium guajava leaves had significantly elevated levels of HSPs genes after challenge with virulent strain of A. hydrophila for 2 weeks (Giri et al 2015). The increase in the HSPs level was attributed to the increased expression and accumulation of the stress proteins which facilitates the ability of cells to both repair and synthesize new proteins to replace those that were damaged after a particular metabolic insult (Ojima et al 2005). The ability of the affected tissue or organ to mount a robust stress response is thought to be important for its survival and recovery (Giri et al 2015). However, the expression of different HSP70 abundance differed in O. niloticus fed 0.5g/kg and 2.5g/kg supplementation levels in treatments MLST5 and MLST25 respectively suggesting a minimum and maximum threshold for the supplementation of Moringa leaf extracts. In the current study, it was found that dietary supplementation with Moringa leaves at concentrations of 1g/kg (MLST10), 1.5g/kg (MLST 15) and 2.5g/kg (MLST20) significantly increased the expression of HSP 70 genes to the control group. Oral administration of similar phytogenic product like Achyranthes aspera leaves augmented the expression of IL-1b and TNF-an immune response genes in Catla catla (Chakrabarti et al , 2014). In present study, the higher post-challenge survival rate of O. niloticus in the MLST10 group was boosted by the immunostimulatory effect of Moringa in stimulating the protective expression of the HSP 70 of the fish in this study. Ojima et al (2005) reported that under conditions of stress, where protein folding/assembly events may be compromised, the increased expression and accumulation of the stress proteins facilitates the ability of fish cells to both repair and synthesize new proteins to replace those that were damaged.
Anwar F Latif S, Ashraf M and Gilani A H 2007 Moringa oleifera : a food plant with multiple medicinal uses. Phytotherapy Research 21(1): 17-25.
AOAC 1995 Association of Official Analytical Chemists Official Methods of Analysis. (16th edition.) Arlington, VA, USA. 1056 pp.
Blancheton J P, Attramadal K J K, Michaud L, d’Orbcastel E R and Vadstein O 2013 Insight into bacterial population in aquaculture systems and its implication. Aquacultural Engineering 53: 30-39.
Chakrabarti R, Srivastava P K, Verma N and Sharma J G 2014 Effect of seeds of Achyranthes aspera on the immune responses and expression of some immune-related genes in carp Catla catla, Fish Shellfish Immunology 4: 164-69.
Conte F S 2004 Stress and the welfare of cultured fish.Applied Animal Behaviour Science 86(3): 205-223.
FAO 2014 Food and Agricultural Organisation of the United Nations: The state of world fisheries and Aquaculture Rome, Italy. 370 pp.
Giri S S, Sen S S, Chi C, Kim H J ,Yun S, Park S C and Sukumaran V 2015 Effect of guava leaves on the growth performance and cytokine gene expression of Labeo rohita and its susceptibility toAeromonas hydrophila infection. Fish and shellfish immunology 46(2): 217-224.
Guo Q ,Liu F, Shi H, Yu Y , Wang Y and Shen W Lv F 2016 Effects of astaxanthin and emodin on the growth, stress resistance and disease resistance of yellow catfish (Pelteobagrus fulvidraco). Fish and Shellfish Immunology, 5: 125-135.
Hammed A M, Amosu A O, Awe A F and Gbadamosi F F 2015 Effects of Moringa oleifera leaf extracts on bacteria (Aeromonas hydrophila) infected adult African mud cat fish.International Journal of Current Research 7 (10): 22117-22122.
Kumar S, Abhay K and Pandey A K 2013 Chemistry and Biological Activities of Flavonoids: An Overview. The Scientific World Journal 13: 1-16.
Lim C and Webster C D 2006 Tilapia: Biology, Culture, and Nutrition. Haworth Press. USA. 357 pp.
Li L, Kobayashi M, Kaneko H, Nakajima-Takagi Y, Nakayama Y and Yamamoto M 2008 Molecular evolution of Keap1: Two Keap1 molecules with distinctive intervening region structures are conserved among fish. Journal of Biological Chemistry 283(6):3248–3255.
Liu Z, Xie Y, Song L, Weng Z and Liu S 2015 Hsp90, Hsp60 and Hsp families of heat shock protein genes in channel catfish and their expression after bacterial infections. Fish and Shellfish Immunology 44: 642-651.
Makanjuola O O, Dada E O and Ekundayo F O 2013 Antibacterial activities of Moringa oleifera on coliforms isolated from some surface waters in Akure, Nigeria.FUTA Journal of Research in Sciences 1: 63-71.
Ojiako E N 2014 Phytochemical analysis and antimicrobial screening ofMoringa Oleifera leaves extract. The International Journal of Engineering and Science 3(3): 32-43.
Ojima N, Yamashita M and Watanabe S 2005 Quantitative mRNA expression profiling of heat shock protein families in rainbow trout and Nile tilapia cells. Biochemical and biophysical resources communication 329:51–57.
Rapatsa M M and Moyo N A G 2014 Effect of Moringa oleifera on the histology, haematology and growth of Oreochromis mossambicus in aquadams® fertilised with chicken manure in South Africa. African Journal of Aquatic Science 39: 295-303.
Sen S S, Giri S S and Sukumaran V 2014 Role of HSP70 in cytoplasm protection against thermal stress in rohu, Labeo rohita. Fish and Shellfish Immunology 41: 294-299.
Tekle W and Sahu, N P 2015 Growth and immunodulatory response of Nile tilapia,Oreochromis niloticus fingerlings to ethanolic extract ofMoringa oleifera flower. International Journal of Sustainable and Renewable Products 5(7): 285-296.
Wang F, Yunliang L, Zhang D and Dong S 2016 Hypothermal effects on survival, energy homeostasis and expression of energy-related genes of swimming crabs Portunus trituberculatus during air exposure. Journal of Thermal Biology 60: 33–40.
Wedemeyer G A, Barton B A. and McLeay D J 2000 Stress and acclimation. In: Methods for Fish biology. (Editors: Schreck, C. and Moyle, P). American Fisheries Society, Bethesda, Maryland 7: 451-489.
Xie J K, Jun S K and Jian F B 2008 Effects of anthraquinone extract from rhubarb, Rheum officinale on the crowding stress response and growth of common carp,Cyprinus carpio var. Jian. Aquaculture 281: 5-11.
Received 15 February 2017; Accepted 6 March 2017; Published 1 April 2017