
2015 compared to the study period of January 2016 to December 2016 for a cold high-elevation steppe near Laramie, WY, USA.
Time series data were derived from ‘parameter-elevation relationships on independent slope model’ (PRISM) interpolation.
| Livestock Research for Rural Development 29 (11) 2017 | Guide for preparation of papers | LRRD Newsletter | Citation of this paper |
Sustainable pastoralism on North American high-elevation cold steppes requires an understanding of forage dynamics relative to potential compositional changes. I sampled forage quality monthly and peak forage mass of a native C4 grass (Bouteloua gracilis), a native C3 grass (Pascopyrum smithii), and an exotic C3 grass (Agropyron cristatum) in 2016, an average precipitation and temperature year, on a 2,195 m elevation steppe grassland in Wyoming USA. Constrained ordination of the forage quality data displayed the first axis by photosynthetic pathways and the second axis by different native/exotic origins (86.3% fit variation explained; permutation test p = 0.003). Forage species, day of year, and their interaction were significant for all forage quality metrics (p < 0.001) indicating that species-specific forage quality characteristics respond to seasonal changes differently. For example, A. cristatum had the earliest crude protein peak in April, followed by P. smithii from May – July, followed by B. gracilis in August. Other examples include lower lignin values for B. gracilis, and P and K peaks for P. smithii. A. cristatum yielded greater forage mass than P. smithii or B. gracilis. Relative to plant succession and potential compositional change, these three species combined match a broader range of livestock nutritional needs than any single species.
Keywords: forage ecology, grassland, grazing, herbage, pastoralism, rangeland
Forage-based livestock production and pastoralism continues to be a dominant feature of cold high-elevation steppes globally. These vast and inhospitable ecosystems occur in central Asia, western North America, southwestern South America, South Africa, southern Australia, and a few other areas. These areas are classified as BSk (B – Arid; S – Steppe; k - cold) based on the Köppen-Geiger climate classification, are temperate, continental, and characterized by snowfall in the winter and large diurnal temperature gradients (Peel et al 2007). The physiognomy of cold high-elevation steppes makes them unsuitable for farming and crop production due to short growing seasons, cold temperatures, low precipitation, high wind speeds, shallow soils, and difficult terrain (Ormaechea and Peri 2015). However, these features have also conserved native floristic features of cold steppes due to the suitability only for extensive pastoral agriculture (Yabrir et al., 2015). Finally, the cold end of the thermal gradient is a dominant and controlling feature of cold steppes. For example, in a Patagonian steppe, climatic controls of annual net primary productivity (ANPP) were strongly controlled by an interaction of temperature and precipitation, and notably, the lower and cooler end of the thermal gradient (Jobbágy et al 2002). Thus, photosynthetic pathway of forage species is likely to interact with such thermal extremes that characterize cold steppes.
Sustainability of pastoral livelihoods and livestock production, and the concomitant conservation of flora and fauna, in steppes should include an enhanced understanding of species-specific forage dynamics in the context of potential change (Dong et al 2011, Maamri et al 2015). Moreover, providing livestock with naturally grown forage continues to be the most economical and efficient means of meeting nutritional requirements (Short 2001, Meena et al 2007). From a short-term animal nutrition perspective, the long and cold winters are periods of no forage plant photosynthetic activity and subsequent low forage quality. During these periods livestock may not be able to obtain minimal nutritional requirements (Van Soest 1994). From a long-term forage based production system perspective in steppes, management of grazing relative to forage species, quantity, and quality can induce compositional changes that will have carrying capacity and animal performance consequences (Yousfi et al 2017). In other words, the loss of the most adaptive and productive native species due to mis-management may have negative livestock production consequences (Holechek et al 2004, Derner et al 2008).
Such plant composition and successional changes to native cold steppe plant communities can result from improver grazing management, anthropogenic invasion pathways, or both (Scasta et al 2015). For example, concern about grazing-induced compositional changes relative to both direct and indirect herbivore selectivity effects, and relative to different plant functional traits in Patagonian steppe, have also been demonstrated with intensive grazing leading to greater dominance by low growing grazing-tolerant species (Cingolani et al 2005). Similar plant composition changes have been documented in the northern Great Plains of North America where long-term grazing at heavy stocking rates induced shifts from C3 mid-grass species to C4 short-grass species (Derner et al 2008). While such shifts, relative to grazing are predictable (Augustine et al 2017), the net forage quality and quantity effects are not as well understood. Climate change effects on plant composition, structure, and function further complicate the sustainability of pastoralism on high-elevation steppes (Ganjurjav et al 2016). As one example, a sensitivity analysis of an Inner Mongolia steppe forecasted potential vulnerability of shifting from a grass-dominated state to a shrub-dominated state with increasing temperature, precipitation, and CO2, or broad declines in herbaceous plant production with decreasing precipitation and increasing temperature and CO2 (Christensen et al 2004).
Considering that the sustainability of pastoralism on North American high-elevation cold steppes requires an understanding of forage dynamics relative to potential compositional changes, and the need for enhanced quantification of the temporal dynamics of potential changes, I sampled forage quality monthly and peak growing season forage mass (Allen et al 2011) of blue grama - a native C4 grass (Bouteloua gracilis (Willd. ex Kunth) Lag. ex Griffiths), western wheatgrass - a native C3 grass (Pascopyrum smithii (Rydb.) Á. Löve), and crested wheatgrass - an exotic C3 grass (Agropyron cristatum ((L.) Gaertn.) in 2016 on a 2,195 m elevation steppe grassland in Wyoming USA. This information is particularly important for pastoralists in Wyoming USA because Wyoming is the least populous state in the USA with an estimated 69% of the state population classified as rural and > 68% of total state farm receipts derived from rangeland cattle production (USDA ERS 2017). My objectives were to (1) determine how forage quality composition of these three species was explained relative to photosynthetic pathway and native/exotic origins, (2) quantify seasonal trends relative to important forage quality characteristics, (3) analyze annual differences between species for important forage quality characteristics, and (4) elucidate value of a diverse forage assemblage for livestock nutritional demands and potential consequences to shifts to homogenous stands of each of these three forage species.
The general study area is owned and managed by the University of Wyoming – Laramie Agriculture Experiment Station and occurs in an ecotone between the northern mixed grass prairie and sagebrush steppe of southeastern, WY, USA. The study site consisted of a 5 ha pasture embedded within the larger 400 ha working research ranch. The study site was not grazed by livestock in 2016, the period of study. Study site soils consist entirely of the 165 Forelle-Diamondville association with 3 to 15% slopes and consisted entirely of the R034XY322WY Loamy (10-14SE) ecological site (USDA NRCS 2016). The soils are characterized by a pH of 7.6, cation-exchange capacity of 10.0 milliequivalents per 100 grams, an available water capacity of 0.15 centimeters of water per centimeter of soil, soil bulk density of 1.28 grams per cubic centimeter, 19.3% clay, 57.7% sand, and 22.9% silt in the 0-15 cm layer. The historical “natural” plant community was dominated by P. smithii (35%), needle-and-thread (Hesperostipa comata (Trin. & Rupr.) Barkworth) (15%), bluebunch wheatgrass (Pseudoroegneria spicata (Pursh) Á. Löve) (10%), big sagebrush ( Artemisia tridentata Nutt. subsp. wyomingensis Beetle & Young) (10%), prairie junegrass (Koeleria macrantha (Ledeb.) Schult.) (5%), Indian ricegrass (Achnatherum hymenoides (Roem. & Schult.) Barkworth) (5%), and muttongrass (Poa fendleriana (Steud.) Vasey) (5%). Total forage dry-weight production on the study site is estimated to be 1,568 kg DM ha-1 in favorable years, 1,232 kg DM ha-1 in normal years, and 672 kg DM ha1 in unfavorable years (USDA NRCS 2016).
For the forage quality analyses, I sampled the three dominant grass forage species on our study site, including B. gracilis, P. smithii, and A. cristatum. Entire plants including leaves, stems and seeds were collected by clipping to the ground surface level and immediately placing plant material in a paper bag. The only criteria used for not sampling was to avoid areas clipped the previous month. Forage samples were of a minimum of 50 grams and were randomly collected from the study site on a monthly basis. Sampling began January 13 th 2016 and continued through December 2016. Samples were always collected at the mid-point of each month. Four replicates of each species were collected each month (n = 4; 12 months) for a total of 48 samples per species for the year-long study. Samples were placed in a forced air oven and dried for 48-hours at 60 °C (Elgersma et al 2014).
For the forage mass analyses, I sampled the same three dominant grass forage species in late-September 2016. The sampling method was designed to estimate potential maximum forage mass of the three plant species. In other words, the sampling method estimates the potential maximum mass of each species if the plant composition on the study site were comprised of a monoculture of that species (as opposed to relative contributions of each species). This protocol was selected because the plant community dynamics relative to either A. cristatum invasion or B. gracilis dominance from grazing could shift the plant community and associated forage quality/quantity characteristics different directions with implications for livestock production. These forage mass samples were collected with a 0.25 m2 quadrat by clipping all of an individual species within the quadrat to the ground surface level and immediately placing plant material in a paper bag. Sampling was stratified across five randomly selected locations within the study site for five replications (n = 5) per plant species. At a random location, I first determined if all three species occurred. Then I positioned the quadrat on a spot dominated by one of the three species and clipped only the plants of that species. The next two species were then collected within 5 m of the first species in different quadrats positioned in the nearest spot dominated by each of the other two species. Samples were placed in a forced air oven and dried for 48-hours at 60 °C (Elgersma et al 2014) and then weighed and converted to kilograms per hectare.
Monthly samples collected for forage quality were sent to Ward Laboratories, Inc. in Kearney, NE, USA for laboratory analysis and all results were reported on a dry (0.00% moisture and 100.00% dry matter) basis. Ward Laboratories is certified by the National Forage Testing Association (Ward Laboratories Inc 2017). Individual samples were ground at the lab to pass through a 1 mm sieve prior to analyses. Near infrared reflectance (NIR) spectrometry was then conducted using a Foss 2500 at wavelengths between 850 and 2500 nm to quantify presence of key forage quality factors based on universal equations generated from a suite of forage species (Marten et al 1989, Schutte and Lauriault 2015). Specifically, current calibration of the NIR technique are based on 2013 NIRS Level 2 equations (NIRS Forage and Feed Testing Consortium 2017). Key forage quality factors assessed included: crude protein (CP), acid detergent fiber (ADF), neutral detergent fiber (NDF), NDF 48-hour digestibility (NDFD), in vitro true 48-hour digestibility (IVTDMD), total digestible nutrients (TDN), net energy for lactation, net energy for maintenance, net energy for gain, relative feed value (RFV), relative forage quality (RFQ), calcium (Ca), phosphorous (P), potassium (K), magnesium (Mg), ash, fat, lignin, non-fiber carbohydrates, and water-soluble carbohydrates. We had sample quantity issues only for B. gracilis in March and because NIRS was not possible, samples were assessed with wet chemistry methods that provide only a subset of forage quality metrics listed above.
In order to account for the complex forage quality data, I first used multivariate statistics by conducting a direct gradient ordination analyses of forage quality response metrics constrained by forage species using redundancy analysis (RDA) which is a constrained form of principal components analysis (PCA), an indirect and unconstrained ordination technique (Gusmeroli et al 2013, Chinea and Arévalo 2014, Hakl et al 2015). The RDA ordination included ‘day of year’ as a covariate to account for the intra-annual effect on forage quality. Response data was log-transformed, centered, and standardized. All constrained axes were tested for significance at α 0.05 using a permutation test with 999 iterations and random seed number generator. Ordination analyses were conducted in CANOCO (Lepš and Šmilauer 2003, Ter Braak and Šmilauer 2012).
To further understand forage quality dynamics, I then conducted analyses of variance (ANOVA) separately for each forage quality metric and graphically displayed them with line graphs and bar graphs in four general categories: quality, energy, fiber and digestibility, and mineral nutrient content. For each ANOVA, I used a mixed effects model with Species, Day of Year, and their interaction (Species*Day) as the main fixed effects. I also assigned Day of Year as a repeated effect because each species was repeatedly sampled at a regular and increasing Day of Year interval. To compare means and determine significant differences I assessed the Type II hypothesis test, and evaluated all pairwise comparisons that were considered different at α 0.05. All forage quality metrics that are percentage data were arcsine square-root transformed to meet the assumptions of normality and distribution for our ANOVA’s but all of the raw means and standard errors are presented in the figures. I did not conduct this analysis for the three net energy values (for lactation, for maintenance, for gain) because these are extrapolations based on TDN values and are highly correlated based on our ordination analyses and estimates will be similar to the TDN relationships and analyses. Similarly, to further understand forage mass differences, I also conducted an ANOVA with a mixed effects model with forage species as the fixed effect and sample replication as a random effect to account for soil, water, and topographic influences at the microsite level. To compare means and determine significant differences I assessed the Type II hypothesis test, and evaluated all pairwise comparisons that were considered different at α 0.05.
Precipitation during our period of study followed the typical seasonal pattern to be expected with the greatest amounts coming in the spring (March, April, May, and June). Late summer and fall also expressed the seasonal decline in precipitation with a drier than normal July and August. It should be noted that these two months were still not the driest on record (Figure 1a). However, precipitation received in August, September, and November offset those lows. Temperature during our period of study also followed the typical seasonal pattern to be expected with the coldest months occurring in the winter (January, February, and December) and the warmest months in the summer (July and August) (Figure 1b).
|
| Figure 1.
Box plots displaying the historical monthly (a) precipitation (mm) and (b)
mean temperature (°C) data distribution from 1895 to 2015 compared to the study period of January 2016 to December 2016 for a cold high-elevation steppe near Laramie, WY, USA. Time series data were derived from ‘parameter-elevation relationships on independent slope model’ (PRISM) interpolation. |
The redundancy analysis (RDA) ordination constrained to the three forage species and day of year revealed that the three forage species have different forage quality composition with unique forage quality parameters for each. The first axis explained 61% of the fitted variation and is largely a function of the differences associated with photosynthetic pathway and growth form – C3 and upright species grouped on the left side and the C4 and low-growing species on the right side of the biplot (Figure 2). Forage quality response variables explaining this difference include in vitro digestibility, ash content, and acid detergent fiber (ADF) content that all increase in association with B. gracilis, versus lignin, Mg, and non-fiber carbohydrates that all increase in association with A. cristatum and P. smithii. The second axis explained 25% of the fitted variation, with the first two axis explaining a cumulative of 86.3% of the fitted variation. The second axis is largely a function of species origin differences associated with the exotic and native C3 species, with A. cristatum at the top of the biplot and P. smithii at the bottom of the biplot, and the associated patterns of these species forage quality composition relative to seasonal changes as demonstrated by the ‘Day of year’ arrow (Figure 2). Forage quality response variables explaining this difference include lignin and neutral detergent fiber (NDF) that increase in association with A. cristatum, versus fat, phosphorous, TDN (and associated net energy content calculations for maintenance, lactation, and gain) and neutral detergent fiber digestibility (NDFD) that all increase in association with P. smithii. Finally, the permutation test for significance of all constrained axes was significant (pseudo-F = 3.2; p = 0.003) indicating the three forage species had different forage quality composition (Figure 2).
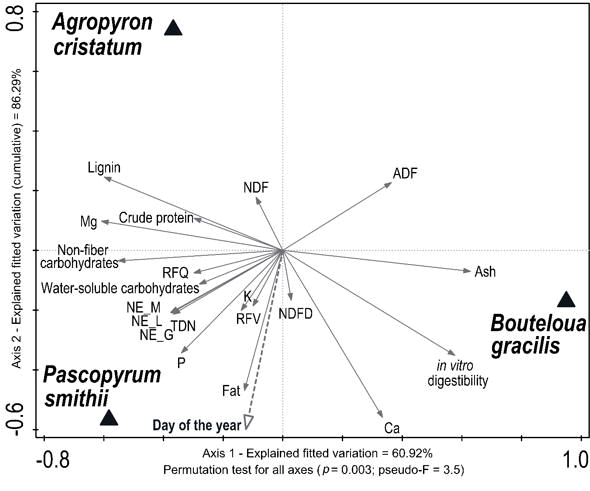 |
| Figure 2.
Ordination diagram of forage quality metrics for three grass species based
on a redundancy analysis (RDA) constrained by Species and Day of Year sampling intervals. Forage quality data was collected monthly for 2016 in a cold high-elevation steppe near Laramie, WY, USA. Significance of constrained axes is based on a Monte Carlo permutation test with 999 iterations. |
For all metrics of forage quality, forage species, day of year, and the species*day interaction were all significant (p < 0.0001) indicating that certain species offer greater quality and respond to seasonal changes differently (Figure 3a-c). Prior to April, crude protein (CP) was below 7% for all species. The first increase in CP was for A. cristatum in April to 9.9 ± 1.0% followed by a subsequent decline the rest of the year. In May, P. smithii CP increased to the highest level and remained the highest of the three species through June and July with a maximum level of 12.2 ± 0.4%. In August, B. gracilis offered the highest CP (9.1 ± 0.3%). Annual mean CP (± standard error) for B. gracilis, P . smithii, and A. cristatum were 4.8 ± 1.6%, 7.1 ± 1.4%, and 5.6 ± 1.2% respectively. Precipitation in November led to an increase in CP for both C3 species in December but not for the C4 species (Figure 1; Figure 3a). Relative forage quality (RFQ) and relative forage value (RFV) followed a similar pattern as expressed by CP (Figure 3b and 3c). P. smithii had the highest RFQ of all species in June and July with values exceeding 125. The C4 species B. gracilis had low RFQ until July when temperatures warmed sufficiently for substantial photosynthetic activity and it reached an average of 125. A. cristatum had the lowest RFQ for 6 months of the year and only exceeded 100 one month (June). RFV followed a similar pattern as RFQ (Figure 3c).
 |
| Figure 3.
Forage quality data expressed as (a) crude protein, (b) relative feed
quality (RFQ), and (c) relative feed value for three grass species sampled monthly for 2016 in a cold high-elevation steppe near Laramie, WY, USA. Grass species include Bouteloua gracilis - a C4 low-growing native, Pascopyrum smithii - a C3 upright native, and Agropyron cristatum - a C3 upright exotic. |
For all metrics of energy, forage species, day of year, and the species*day interaction were all significant (p < 0.0001) indicating that certain species offer greater energy and respond to seasonal changes differently (Figure 4a-c). Differences in non-fiber carbohydrates was variable with the C3 species (P. smithii and A. cristatum ) generally displaying higher values than the C4 species (B. gracilis) (Figure 4a). Differences in water-soluble carbohydrates were not apparent until May when A. cristatum offered the greatest content which also occurred in July, August, and September (Figure 4b). Fat content followed a pattern similar to TDN with B. gracilis displaying higher values early in the year, P. smithii displaying higher values later in the year and the only species to exceed an average of 4% in any given month, and A. cristatum consistently displaying the lowest values (Figure 4d). Total digestible nutrients (TDN) were variable until June when P. smithii offered the greatest TDN content from then until the end of the year (Figure 4d). B. gracilis offered the lowest TDN content four months of the year and was similar to A. cristatum seven months of the year.
 |
| Figure 4.
Forage energy data expressed as (a) non-fiber carbohydrates, (b)
water-soluble carbohydrates, (c) fat, and (d) total digestible nutrients
(TDN), for three grass species sampled monthly for 2016 in a cold high-elevation steppe near Laramie, WY, USA. Grass species include Bouteloua gracilis - a C4 low-growing native, Pascopyrum smithii - a C3 upright native, and Agropyron cristatum - a C3 upright exotic. |
For all metrics of fiber and digestibility, forage species, day of year, and the species*day interaction were all significant (p < 0.0001) indicating that certain species have greater fiber content or lower digestibility and respond to seasonal changes differently (Figure 5a-c). Lignin content was highest in A. cristatum, intermediate in P. smithii, and lowest in B. gracilis, with a greater separation later in the year (Figure 5a). Acid detergent fiber (ADF) tended to be lower in P. smithii for at least half of the year than the other two species and B. gracilis had the highest ADF in February, April, May, and December (Figure 5b). Neutral detergent fiber (NDF) was variable throughout the year with each forage species having the highest NDF at least one month of the year and A. cristatum having the highest NDF in January, June, August, October, November, and December (Figure 5c). The inverse of the lignin and species temporal relationship was reflected similarly in the pattern of NDF 48-hour digestibility (NDFD) and in vitro true 48-hour digestibility (IVTDMD) (Figure 5d and 5e). B. gracilis had the highest levels of NDFD and IVTDMD, P. smithii had intermediate levels of NDFD and IVTDMD, and A. cristatum had the lowest levels of NDFD and IVTDMD with the exception of May (Figure 5d and 5d).
 |
| Figure 5.
Forage fiber and digestibility data expressed as (a) lignin, (b) acid
detergent fiber (ADF), (c) neutral detergent fiber (NDF), (d) neutral detergent fiber digestibility (NDFD), and (e) in vitro dry matter digestibility for three grass species sampled monthly for 2016 in a cold high-elevation steppe near Laramie, WY, USA. Grass species include Bouteloua gracilis - a C4 low-growing native, Pascopyrum smithii - a C3 upright native, and Agropyron cristatum - a C3 upright exotic. |
For all metrics of mineral nutrient content, forage species, day of year, and the species*day interaction were all significant (p < 0.0001) indicating that certain species have specific mineral content and respond to seasonal changes differently (Figure 6a-d). Ash content was highest in B. gracilis (12.0 ± 2.1%) and lowest in A. cristatum (3.0 ± 1.0%) and P. smithii (3.3 ± 1.1%) (Figure 6a). Calcium (Ca) content was highest in B. gracilis (0.41 ± 0.06%), intermediate in P. smithii (0.34 ± 0.06%), and lowest in A. cristatum (0.26 ± 0.05%) with a similar Ca low in the May sampling period for all three species (Figure 6b). Magnesium (Mg) content was the highest in P. smithii, (0.13 ± 0.01%), intermediate in A. cristatum (0.11 ± 0.17%), and lowest in B. gracilis (0.07 ± 0.02%) (Figure 6c). Phosphorous (P) content was higher in P. smithii (0.12 ± 0.03%) than B. gracilis (0.06 ± 0.02%) or A. cristatum (0.07 ± 0.01%), and the seasonal expression of this difference was apparent in growing season only when P. smithii had higher values (Figure 6d). Similarly, potassium (K) content was higher in P. smithii (0.46 ± 0.25%) than B. gracilis (0.27 ± 0.14%) or A. cristatum (0.22 ± 0.13%), and the seasonal expression of this difference was apparent in growing season only when P. smithii had higher values (Figure 6e).
 |
| Figure 6.
Forage mineral nutrient content data expressed as (a) ash, (b) calcium (Ca),
(c) magnesium (Mg), (d) phosphorous (P), and (e) potassium (K) for three grass species sampled monthly for 2016 in a cold high-elevation steppe near Laramie, WY, USA. Grass species include Bouteloua gracilis - a C4 low-growing native, Pascopyrum smithii - a C3 upright native, and Agropyron cristatum - a C3 upright exotic. |
B. gracilis forage mass potential did not display inter-species sample differences (p = 0.08; Figure 7a) with samples ranging from 188 to 448 kg DM per ha. However, P. smithii and A. cristatum forage mass potential both displayed inter-species sample differences (p < 0.05 respectively) with some samples higher than others (Figure 7b and 7c). P. smithii samples ranged from 480 to 2,064 kg DM per ha and A. cristatum samples ranged from 712 to 3,940 kg DM per ha. Comparison of mean species forage mass potential was significant (P < 0.05) with A. cristatum yielding greater forage mass (2,012 ± 560 kg DM per ha) than P. smithii (969 ± 284 kg DM per ha), and both A. cristatum and P. smithii yielding greater forage mass than B. gracilis (325 ± 48 kg DM per ha) (Figure 7d).
 |
| Figure 7.
Potential maximum forage mass for three grass species sampled monthly for
2016 in a cold high-elevation steppe near Laramie, WY, USA. Grass species include Bouteloua gracilis - a C4 low-growing native, Pascopyrum smithii - a C3 upright native, and Agropyron cristatum - a C3 upright exotic. |
The diverse forage quality and quantity characteristics of the three forage species analyzed in our study indicates that a broader range of livestock nutrition benefits can be provided collectively better than any single species can provide singularly (Provenza et al 2009). The combination of these three species is most beneficial in crude protein by initiating earlier and extending later the optimal crude protein and by only certain species offering adequate levels of certain nutrients during the growing season (i.e., P. smithii and P and K). However, these results must be considered within the context of plant succession because the current plant community is different from the historical “natural” plant community. Currently, A. cristatum and B. gracilis now comprise a substantial proportion of the plant community on the study site. We hypothesize these species have established by different mechanisms. First, A. cristatum is present in the region due to its introduction as an alternative forage from Asia and has established at our study site from the movement of hay and vehicles and/or dispersal of seed by other means (Scasta et al., 2015). Second, B. gracilis has established from the ecological site immediately to the north (151 Diomandville-Cushool complex with 3 to 15% slopes) where it was a minor component of the plant community and is a consequence of a history of livestock grazing that has also eliminated or reduced perennial cool-season grasses (Derner et al 2008, USDA NRCS 2016). Finally, one additional contemporary difference is the absence of big sagebrush (Artemisia species) on the study site, and generally within the Laramie Valley. This is why the area is characterized as a high-elevation steppe rather than a sagebrush steppe.
Our results also demonstrate the compatibility of these three forage species for optimizing livestock nutritional resources in this North American steppe. Implications for successional changes for forage-based livestock productions systems if the plant community approached a monoculture for either of the three forage species assessed also becomes apparent. First, if P. smithii was the primary forage species as was historically documented, total forage mass and carrying capacity would be 3x greater than if B. gracilis dominated and ½ of the potential if A. cristatum dominated but crude protein, total energy (TDN), forage quality (RFQ and RFV), fat, P, and K would be optimized on an annual basis. However, the early-season crude protein in A. cristatum (March) and late-season crude protein in B. gracilis (August) would be lost. Second, if B. gracilis was the primary forage due to the effects of grazing and loss of the C3 grasses (Cingolani et al 2005, Derner et al 2008), total forage mass and carrying capacity would be 1/3 rd to 1/6th lower and early season forage quality below animal nutrient requirements until June due to the cold environment relative to the C4 photosynthetic pathway. However, B. gracilis coupled with the other two forage species does provide an August crude protein extension, is the optimal species in terms of lignin content and digestibility (NDFD and in vitro), and the optimal source of Ca and ash. Finally, if A. cristatum was the primary forage due to continued invasion and selection for this species due to its lower digestibility, total forage mass and carrying capacity would be 2x to 6x greater but crude protein would be lower, fat would be lowest, and is the worst species in terms of lignin content, digestibility ((NDFD and in vitro), and Ca. The presence of A. cristatum in native steppe has been documented to produce 2x total forage mass by Hart et al (1983b) which also translated into greater pounds of calf production per unit area. However, A. cristatum monocultures are unlikely to meet livestock nutritional needs in the late-summer and fall, especially if moisture is limiting (Angell et al 1990) – an issue that our results also demonstrate.
Because the livelihood of cold steppe pastoralists depends upon the reproduction of ruminant livestock such as sheep and goats and cecal fermenters such as horses, we placed our results in the context of critical nutrients requirements at reproductive stages for each livestock species. First, I consider nutrient requirements for average milking (5 kg milk/day) beef cows of moderate size (450 kg) in the first 3 to 4 months postpartum are 57.5% TDN, 9.9% CP, 0.28% Ca, and 0.23% P (National Research Council 1984, Cheeke 1991). For such cows, the highest TDN values offered by P. smithii 6 months of the year are critical, the early boost of CP offered by A. cristatum is important for early calving cows and the late boost of CP offered by B. gracilis is important for later calving cows, the Ca offered by B. gracilis during March and May are critical, and of the three forage species only P. smithii meets the minimum P requirement for at least a single month of the year. I can make similar conclusions regarding sheep when the average nutrient requirements for 70 kg ewes in the last 4 weeks of gestation with an expected lambing rate of 130-150% are 59% TDN, 10.7% CP, 0.35% Ca, and 0.23% P and the average nutrient requirements for these same ewes in the first 6-8 weeks of lactation with twin lambs are 65% TDN, 15.0% CP, 0.39% Ca, and 0.29% P (National Research Council 1985, Cheeke 1991). In this scenario, minimum nutrient requirements are even higher than for beef cows, and thus, the forage diversity has similar direct animal production and nutrient requirement implications. Finally, for horses the average nutrient requirements for lactating mares from foaling to 3 months are 13.2% CP, 0.52% Ca, 0.34% P, 0.10% Mg, and 0.42% K (National Research Council 1989, Cheeke 1991). As for beef cows and sheep, different species meet different minimum nutrient requirements at different times and during the summer P. smithii is absolutely critical for meeting minimum Mg and K requirements that B. gracilis and A. cristatum do not meet.
Diverse forage quality and quantity characteristics are critical for sustainable pastoralism (Yousfi et al 2017) on our high elevation steppe study area and are also indicators of species richness and diversity. Moreover, the harsh extremes of the high-elevation, cold, and semi-arid steppe environment of our study suggests that forage species must be well adapted for local climates. Our results however indicate that local adaptation is not important simply at the regional level but also at the temporal level, and specifically at the intra-annual scale. In other words, each of the three species demonstrates a forage quality optima at different times of the year. For A. cristatum, this is earlier in the season, for P. smithii this is mid-season, and for B. gracilis this is late-season as expressed by the respective initiation of photosynthetic activity and crude protein values. The forage quality and quantity features of each forage species also results in tradeoffs that pastoralists will need to be made aware, especially if they are considering conversion to an A. cristatum monoculture or are grazing at an intensity that may be leading to successional changes to a B. gracilis dominated plant community. Although grazing induced successional changes are a concern for grazing cold high-elevation steppes, these changes occur with a high level of predictability relative to grazing pressure (Augustine et al 2017). Separate from grazing, the biomass allocation patterns of A. cristatum in steppe environments are shown to be greater for reproductive shoots and seed production which could lead to spread (Wang 2005). Any conversion to A. cristatum monocultures may enhance total forage mass and provide earlier green up and grazing, yet these advantages are offset by the rapid decline in the summer that is more rapid than any other species (Hart et al 1983a). Conceptually and practically, maintaining diverse forage species is critical for sustaining forage-based livestock systems and native wildlife in this high-elevation steppe (Larter and Gates 1991).
This project was supported in part by the University of Wyoming (UW) Department of Ecosystem Science and Management and College of Agriculture and Natural Resources through a USDA National Institute of Food and Agriculture McIntire Stennis Project – ‘Animal-plant interaction ecology on Wyoming Rangelands’ (2015-2020, Project# WYO-559-15). Appreciation is extended to the UW Agricultural Experiment Station staff. Author has no conflict of interest to declare.
Allen V G, Batello C, Berretta E J, Hodgson J, Kothmann M, Li X, McIvor J, Milne J, Morris C, Peeters A and Sanderson M 2011 An international terminology for grazing lands and grazing animals. Grass and Forage Science, Volume 66: 2-28 Retrieved July 1, 2017, from http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2494.2010.00780.x/full
Angell R F, Miller R F and Haferkamp M R 1990 Variability of crude protein in crested wheatgrass at defined stages of phenology. Journal of Range Management, Volume 43, Volume 3: 186-189 Retrieved July 1, 2017, from https://journals.uair.arizona.edu/index.php/jrm/article/download/8470/8082
Augustine D J, Derner J D, Milchunas D, Blumenthal D and Porensky L M 2017 Grazing moderates increases in C3 grass abundance over seven decades across a soil texture gradient in shortgrass steppe. Journal of Vegetation Science, Volume 28, Issue 3: 562-572.
Cheeke P R 1991 Applied Animal Nutrition: Feeds and Feeding. Macmillan, New York, USA.
Chinea E and Arévalo J R 2014 Effects of fertilization management on pasture productivity and nutrient composition. Grass and Forage Science, Volume 69, Issue 3: 415-424.
Christensen L, Coughenour M B, Ellis J E and Chen Z Z 2004 Vulnerability of the Asian typical steppe to grazing and climate change. Climatic Change, Volume 63, Issue 3: 351-368 Retrieved July 1, 2017, from http://www.nrel.colostate.edu/assets/nrel_files/labs/coughenour-lab/pubs/Publications/christensen_climatic_change_04.pdf
Cingolani A M, Posse G and Collantes M B 2005 Plant functional traits, herbivore selectivity and response to sheep grazing in Patagonian steppe grasslands. Journal of Applied Ecology, Volume 42: 50-59 Retrieved July 1, 2017, from http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2664.2004.00978.x/epdf
Derner J D, Hart R H, Smith M A and Waggoner J W 2008 Long-term cattle gain responses to stocking rate and grazing systems in northern mixed-grass prairie. Livestock Science, Volume 117: 60-69.
Dong S, Wen L, Liu S, Zhang X, Lassoie J P, Yi S, Li X, Li J and Li Y 2011 Vulnerability of worldwide pastoralism to global changes and interdisciplinary strategies for sustainable pastoralism. Ecology and Society, Volume 16, Issue 2, Article #10 Retrieved July 1, 2017, from https://www.ecologyandsociety.org/vol16/iss2/art10/ES-2011-4093.pdf
Elgersma A, Søegaard K and Jensen S K 2014 Herbage dry‐matter production and forage quality of three legumes and four non‐leguminous forbs grown in single‐species stands. Grass and Forage Science, Volume 69, Issue 4: 705-716.
Ganjurjav H, Gao Q, Gornish E S, Schwartz M W, Liang Y, Cao X, Zhang W, Zhang Y, Li W, Wan Y, Li Y, Danjiu L, Guo H and Li Y 2016 Differential response of alpine steppe and alpine meadow to climate warming in the central Qinghai–Tibetan Plateau. Agricultural and Forest Meteorology, Volume 223: 233-240.
Gusmeroli F, Della Marianna G, Fava F, Monteiro A, Bocchi S and Parolo G 2013 Effects of ecological, landscape and management factors on plant species composition, biodiversity and forage value in Alpine meadows. Grass and Forage Science, Volume 68, Issue 3: 437-447.
Hakl J, Fuksa P, Konečná J and Šantrůček J 2015 Differences in the crude protein fractions of lucerne leaves and stems under different stand structures. Grass and Forage Science, Volume 71, Issue 3: 413-423.
Hart R H, Abdalla O M, Clark D H, Marshall M B, Hamid M H, Hager J A and Waggoner J W 1983a Quality of forage and cattle diets on the Wyoming High Plains. Journal of Range Management, Volume 36: 46-51.
Hart R H, Waggoner J W, Clark D H, Kaltenbach C C, Hager J A and Marshall M B 1983b Beef cattle performance on crested wheatgrass plus native range vs. native range alone. Journal of Range Management, Volume 36: 38-40 Retrieved July 1, 2017, from https://journals.uair.arizona.edu/index.php/jrm/article/view/7461/7073
Holechek J L, Pieper R D and Herbel C H 2004 Range Management: Principles and Practices., 5th ed. Pearson Prentice Hall, New Jersey, USA.
Jobbágy E G, Sala O E and Paruelo J M 2002 Patterns and controls of primary production in the Patagonian steppe: a remote sensing approach. Ecology, Volume 83, Issue 2: 307-319.
Larter N C and Gates C C 1991 Diet and habitat selection of wood bison in relation to seasonal changes in forage quantity and quality. Canadian Journal of Zoology, Volume 69, Issue 10: 2677-2685.
Lepš J and Šmilauer P 2003 Multivariate Analysis of Ecological Data Using CANOCO. Cambridge University Press, Cambridge, UK.
Maamri F, Arbouche F, Harek D, Zermane N and Alane F 2015 Prediction of digestibility of some pastoral resources from the Algerian steppe. Livestock Research for Rural Development, Volume 27, Issue 5, Article # 93 Retrieved July 1, 2017, from http://www.lrrd.org/lrrd27/5/maam27093.html
Marten G C, Shenk J S and Barton F E 1989 Near Infrared Reflectance Spectroscopy (NIRS): Analysis of Forage Quality. U.S. Department of Agriculture, Agriculture Handbook No. 643. Springfield, Va: National Technical Information Service. 110 p.
Meena H R, Ram H, Singh S K, Mahapatra R K, Sahoo A and Rasool T J 2007 Animal husbandry practices at high altitude (> 6000 feet) in Kumaon region of Uttrakhand, India. Livestock Research for Rural Development, Volume 19, Issue 11, Article# 163 Retrieved July 1, 2017, from http://www.lrrd.org/lrrd19/11/meen19163.htm
National Research Council 1984 Nutrient Requirements of Beef Cattle. National Academies Press. Washington, D.C., USA.
National Research Council 1985 Nutrient Requirements of Sheep. National Academies Press. Washington, D.C., USA.
National Research Council 1989 Nutrient Requirements of Horses. National Academies Press. Washington, D.C., USA.
NIRS Forage and Feed Testing Consortium 2017 NIRS Consortium Level 2 Equation Release Statistics 2013 Retrieved March 31, 2017, from http://nirsconsortium.org/Resources/Documents/2013EquationReport-Level2.pdf
Ormaechea S and P Peri 2015 Landscape heterogeneity influences on sheep habits under extensive grazing management in Southern Patagonia. Livestock Research for Rural Development, Volume 27, Issue 6, Article #105 Retrieved July 1, 2017, from http://www.lrrd.org/lrrd27/6/orma27105.html
Peel M C, Finlayson B L and McMahon T A 2007 Updated world map of the Köppen-Geiger climate classification. Hydrology and Earth System Sciences Discussions, Volume 4, Issue 2, 439-473 Retrieved July 1, 2017, from http://www.hydrol-earth-syst-sci.net/11/1633/2007/hess-11-1633-2007.pdf
Provenza F D, Villalba J J, Wiedmeier R W, Lyman T, Owens J, Lisonbee L, Clemensen A, Welch K D, Garder D R and Lee S 2009 Value of plant diversity for diet mixing and sequencing in herbivores. Rangelands, Volume 31: 45-49.
Scasta J D, Engle D M, Fuhlendorf S D, Redfearn D D and Bidwell T G 2015 Meta-analysis of exotic forages as invasive plants in complex multi-functioning landscapes. Invasive Plant Science and Management, Volume 8, Issue 3: 292-306 Retrieved July 1, 2017, from http://www.bioone.org/doi/pdf/10.1614/IPSM-D-14-00076.1
Schutte B J and Lauriault L 2015 Nutritive value of field bindweed (Convolvulus arvensis) roots as a potential livestock feed and the effect of Aceria malherbae on root components. Weed Technology, Volume 29, Issue 2: 329-334.
Short S D 2001 Characteristics and Production Costs of U.S. Cow-Calf Operations. Statistical Bulletin Number 974-3. United States Department of Agriculture, Economic Research Service, Retrieved July 1, 2017, from https://www.ers.usda.gov/webdocs/publications/47150/16084_sb974-3_1_.pdf?v=41063
Ter Braak C J F and Šmilauer P 2012 Canoco Reference Manual and User’s Guide: Software for Ordination, v. 5.0. Microcomputer Power. Ithaca, NY, USA.
USDA ERS 2017 Wyoming Data for Rural Analysis. Economic Research Service, United States Department of Agriculture, Retrieved June 29, 2017, from https://data.ers.usda.gov/reports.aspx?StateFIPS=56&StateName=Wyoming&ID=17854
USDA NRCS 2016 Web Soil Survey. Soil Survey Staff, Natural Resources Conservation Service, United States Department of Agriculture.
Van Soest P J 1994 Nutritional ecology of the ruminant. Cornell University Press.
Ward Laboratories Inc 2017 Certifications/Proficiency, Retrieved March 31, 2017, from https://www.wardlab.com/about.php
Wang R Z 2005 Demographic variation and biomass allocation of Agropyron cristatum grown on steppe and dune sites in the Hunshandake Desert, North China. Grass and Forage Science, Volume 60, Issue 1: 99-102.
Yabrir B, Laoun A, Chenouf N S and A Mati 2015 Characteristics of sheep farms in middle Algerian area steppe in relationship with the aridity of the environment: case of the region of Djelfa. Livestock Research for Rural Development, Volume 27, Issue 10, Article #207 Retrieved July 1, 2017, from http://www.lrrd.org/lrrd27/10/yabr27207.html
Yousfi M, Chachoua I and Bougouffa N 2017 The rangelands contribution in feeding small ruminants in steppe and Djelfa wilaya, Algeria. Livestock Research for Rural Development, Volume 29, Issue 2, Article #39 Retrieved July 1, 2017, from http://www.lrrd.org/lrrd29/2/moha29039.html
Received 6 July 2017; Accepted 10 October 2017; Published 2 November 2017