 |
| Figure 1. Position of AG (AG-CC) to (GG-CC) transition in exon 3 of the gene at 727 base (accession no Genbank GQ452268, Gul et al 2009), or at 255 base of PCR product which alters a HaeIII restriction site |
| Livestock Research for Rural Development 26 (6) 2014 | Guide for preparation of papers | LRRD Newsletter | Citation of this paper |
The objective of the study was to examine whether GH|HaeIII gen in Indonesia fat tailed sheep (Donggala and East Java sheep breeds) is polymorphic and to examine whether the GH|HaeIII polymorphism associate to their birth weight, growth rate and weaning weight. Seventy genomic DNAs that consisted of 42 Donggala and 28 East Java sheep were isolated from blood sample of those animals. A fragment of GH gene, comprising of a part of intron 2, complete exon 3, complete intron 3, complete exon 4, and a part of intron 4, was amplified. The amplified product with the length of 934 bp was digested with HaeIII restriction enzymes and showed the presence of GH HaeIII polymorphism with three HaeIII genotypes (AA, BB and AB). Bands with ten restricted fragments of 277, 202, 110, 100, 94, 68, 49, 22, 8 and 4 bp were identified as genotype AA. Bands with eleven restricted fragments of 256, 202, 110, 100, 94, 68, 49, 22, 21, 8 and 4 bp were identified as genotype BB. Bands with twelve restricted fragments of 277, 256, 202, 110, 100, 94, 68, 49, 22, 21, 8 and 4 bp were genotyped with AB.
Gene sequencing showed that the difference between allele A and allele B occurred was due to AG (AG-CC) to (GG-CC) transition in exon 3 of the gene at 727 base (accession no Genbank GQ452268, Gul et al 2009), or at 255 base of PCR product which results in that the site is recognized by HaeIII enzyme. Thus, the fragment of 277 was split to two fragments of 256 and 21 bp respectively. In Donggala sheep, AA and AB genotypes had the same genotype frequency of 0.357, whereas BB genotype was 0.286. In East Java sheep AA genotype is the most frequent genotype (0.464), whereas BB and AB were 0.286 and 0.250, respectively. The allele frequencies of A and B in Donggala sheep were 0.536 and 0.464, respectively, whereas those for East Java sheep were 0.589 and 0.411. The degree of heterozygosity of the GH|HaeIII polymorphismsin Donggala and East Java sheep are 0.503 and 0.493, respectively. GH|HaeIII polymorphism affects growth rate and weaning weight both in Donggala and East Java sheep. In Donggala sheep, AA was observed to have significantly higher growth rate than BB. AB, as a heterozygote, did not show any significant differences in growth rate compared to both homozygotes (AA and BB). In East Java sheep, AB was noticed to have significantly higher and lower growth rates than BB and AA, respectively. The pattern of relationship between GH|HaeIII polymorphism and weaning weight seems to be similar to those and growth rate. Genotype AA showed significantly higher weaning weight than genotypes BB both in Donggala (11.6 kg vs 9.68 kg) and East Java (10.83 kg vs 9.37 kg) sheep. Genotype AB did not show significant differences in weaning weight compared to genotypes AA and BB both in Donggala and East Java sheep.
The present study concluded that GH|HaeIII polymorphism could be a genetic marker in sheep selection program for improvement of growth rate and weaning weight in Indonesia fat tailed sheep.
Key words: birth weight, Donggala sheep, East Java sheep, HaeIII enzyme, heterozygosity, growth rate, weaning weight
Growth hormone (GH) is an anabolic hormone synthesized and secreted by the somatotroph cells of the anterior lobe of the pituitary in a circadian and pulsatile manner (Ayuk and Sheppard 2006). GH plays an important role in body growth and metabolism through protein synthesis, protein deposition and fat catabolism in tissues and organs (Gluckman et al 1987), thus GH leads to increased nitrogen retention (Hart and Johnson 1986), decreased energy retained as fat (Eisemann et al 1986), gluconeogenesis and cell division (Neathery et al 1991). GH increases intestinal calcium absorption, thereby enhancing overall bone growth and stimulating chondrocyte proliferation (Boyd and Bauman 1989). GH is a polypeptide hormone of about 22kDa molecular weight, composed of 190 or 191 amino acids.
Donggala and East Java sheep are native breeds of Indonesia and classified as fat-tailed sheep (Mason 1980). One of the outstanding characteristics of these sheep is their ability to adapt to hot and humid environmental conditions as well as extraordinary capacity to utilize feed resources (Husain 2001 and Noor et al 2001). There are about 3,000 head of Donggala sheep in Central Sulawesi and around 1,000,000 head of East Java sheep in East Java of Indonesia (Statistics Indonesia of The Republic Indonesia 2012). The distinguished differences between Donggala and East Java sheep are in color of coat, shape of the tail and presence of horns. Donggala sheep have several combinations in color of coat (brown, black and white) (Husain et al 1992, Malewa 2009), whereas East Java sheep have uniform coat color of white (Noor et al 2001). Donggala sheep have horns in ram and are hornless in females (Malewa 2009) whereas East Java sheep are hornless in both males and females (Noor et al 2001). East Java sheep have a wider and longer tail than Donggala sheep (Malewa 2009).
In ovine breeds, GH polymorphisms have being extensively studied. Tahmoorespur et al (2011) and Tahmoorespur and Ahmadi (2012) have used the PCR-SSCP analysis to investigate GH gen polymorphism in Baluchi sheep and found three SSCP patterns and a significant association between SSCP polymorphisms at the GH gene and birth weight. Hajihosseinlo et al (2013) found five SSCP patterns of GH gene in Makooei sheep and significant relationship between the SSCP polymorphisms and biometric traits. Moradian et al (2013) also found five SSCP pattern of GH gene in Makooei sheep and reported the presence of significant association between the SSCP polymorphisms at GH gene to birth weight, weight at 2, 6, and 9 months and weaning weight.
To our knowledge, no study on growth hormone polymorphism using PCR-RLFP in sheep that is associated to their growth traits was published.
Nowdays, there is an increasing interest in the genetic improvement for the economic traits in Donggala and East Java sheep. The presence of information on association of polymorphic gene with the economic traits will help the breeders to carry out selection program for the economic traits on base of genetic marker. The objective of the research was, therefore, to examine whether GH gene in Donggala and East Java sheep is polymorphic, and to examine whether the RLFP polymorphisms at the GH gene in each breed associate to their growth traits.
The present study was conducted on a total of 100 animals belonging to two Indonesian fat tailed sheep breeds, consisted of sixty head of Donggala sheep reared traditionally by the farmers in Palu Town, Central Sulawesi, Indonesia and forty head of East Java sheep maintained at Breeding Research Station, Jember, East Java, Indonesia.
The lambs were weighed weekly and the growth rate was calculated using birth weight and 3-month weight values. Blood samples were collected from jugular vein around neck of lamb by vacutainer needle into 6 ml tubes containing 1 ml of 10% EDTA. The samples were then stored in cool box containing ice jelly pack during collection process then moved in to the refrigerator (4oC) until ready for DNA isolation.
Seventy of genomic
DNAs that consisted of 42 Donggala and 28 East Java sheep were successfully
isolated from the blood sample using DNA extraction kit (Norgen’s blood genomic
DNA isolation kit, Biotek & Corporation, Canada) according to the manufacturer's
instructions. Polymerase Chain Reaction (PCR) was carried out using Eppendorf
thermocycler (Biometra, Germany) and PCR Green Master Mix (Promega Corporation,
USA) containing bacterially derived Taq DNA polymerase, dNTPs, MgCl2 and
reaction buffers at optimal concentrations for efficient amplification of DNA
templates by PCR. Each reaction mixture consisted of 5 µl of go taq green PCR
master mix, 1 μl of the DNA solution (50 to 100 ng/μl), 1 μl of each primer (5
pmol/μl) and 2 µl deionized water (DdH2O). Amplification for a 934 bp
fragment from the intron II to the intron IV of the oGH gene of Lohi breed
(accession no Genbank GQ452268, Gul et al 2009) was carried out using primer as
follows.
GH-F: 5’-GGAGGCAGGAAGGGATGAA-3’ and GH-R: 5’-CCAAGGGAGGGAGAGACAGA-3’ (Kuulasma 2002).
The amplification was carried out using thermalcycler (Eppendorf Mastercycler) with the following conditions: initial denaturation at 95 °C for 5 min, by 33 cycles of denaturation at 95 °C for 45s; annealing at 60 °C for 45s and extension at 72°C for 45s followed by a final extension at 72 °C for 10 min. Each amplification product was analyzed by electrophoresis on a 2% (w/v) agarose gel, using ethidium bromide staining.
RFLP analysis was conducted to detect polymorphism sites. The PCR amplicons were digested with HaeIII restriction endonuclease (BioLabs Inc, New England) The amount of digestion mixture was 4 µl DDH2O, 1 µl Digest buffer, 1 µl HaeIII restriction enzyme, and 4 µl PCR DNA product. The reaction mixture was digested in a water batch at 37 oC for 4 h and moved into an oven at 70 oC for 15 minutes. Four µl of samples were mixed with 2 µl of loading dye and then samples detected by 6 % polyacrilamide gel electrophoresis (PAGE) and subsequent silver staining.
The electrophoresis product restricted by HaeIII enzyme was interpreted by comparing them with the marker band of 10000 bp. Bands with specific number of fragment and/or length of fragment were identified as a HaeIII genotype.
Genotype frequency represents the ratio of a HaeIII genotype to total population. Mathematics model of genotype frequency (Nei and Kumar 2000) are represented as follows:
Xi = (Gi/N) x 100%
where :
Xi = ith genotype frequency
Gi = number sample of ith genotype
N = total sample
Allele frequency is a ratio of an allele to the overall allele at a locus in the population. Mathematics model for allele frequency (Nei and Kumar 2000) are represented as follows:
Xi= (2nii + ⅀nij)/2N
where :
Xi = ith allele frequency
nii = number sample of ii genotype
nij = number sample of ij genotype
N = total sample
Gene diversity or degree of heterozygosity was calculated according to the formula of Nei and Kumar (2000), as follows.
h = 2n (1 - ∑xi2) / (2n - 1)
where:
h = degree of heterozygosity
xi = ith allele frequency
n = total number of individuals
Data on birth weight, growth rate and weaning weight in each breed were analyzed by a one-way analysis of variance using the Minitab 14 statistical program (Minitab 2003) with mathematical model as follows (Steel and Torrie 1993).
Yij = µ + Gi+εj
where :
Yij = growth traits (birth weight, growth rate or weaning weight)
µ = the overall mean,
Gi = the random effect of the ith HaeIII genotype (i= 3, AA, AB and BB)
εij = the random residual error.
All data were expressed in terms of means.
PCR product or amplicon of Indonesia fat tailed sheep (Donggala and East Java breed) that sized 934 bp is based on gene sequencing of the oGH gene of Lohi breed that are located from the intron II to the intron IV (accession no Genbank GQ452268, Gul et al 2009). The success rate of amplication of 70% indicates that gene sequencing of the oGH gene of Lohi breed had similarity to Indonesia fat tailed sheep.
Allele A had ten fragments, namely 277, 202, 110, 100, 94, 68, 49, 22, 8 and 4 bp, whereas allele B had eleven fragments, namely 256, 202, 110, 100, 94, 68, 49, 22, 21, 8 and 4 bp. Gene sequencing showed that the polymorphism occurred was due to AG (AG-CC) to (GG-CC) transition in exon 3 of the gene at 727 base (accession no Genbank GQ452268, Gul et al 2009), or at 255 base of PCR product which results in that the site is recognized by HaeIII enzyme (Figure 1). Thus, the fragment of 277 was split to two fragments of 256 and 21 bp respectively.
|
| Figure 1. Position of AG (AG-CC) to (GG-CC) transition in exon 3 of the gene at 727 base (accession no Genbank GQ452268, Gul et al 2009), or at 255 base of PCR product which alters a HaeIII restriction site |
Bands with ten restricted fragments of 277, 202, 110, 100, 94, 68, 49, 22, 8 and 4 bp are identified as genotype AA. Bands with eleven restricted fragments of 256, 202, 110, 100, 94, 68, 49, 22, 21, 8 and 4 bp were identified as genotype BB. Bands with twelve restricted fragments of 277, 256, 202, 110, 100, 94, 68, 49, 22, 21, 8 and 4 bp were genotyped with AB.
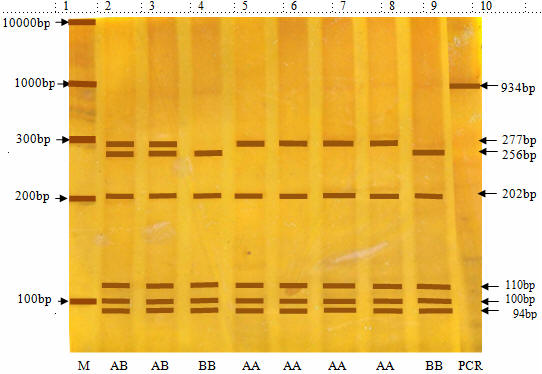 |
Figure 2. Representative genotyping of GH|HaeIII polymorphisms by agarose gel electrophoresis. Strands with 277 bp, 202 bp, 110 bp, 100 bp, 94 bp, 68 bp, 49 bp, 22 bp, 8 bp, 4 bp were genotyped as AA (lanes 5,6,7 and 8); |
In Indonesian fat tailed sheep, whether Donggala or East Java breeds, three GH|HaeIII genotypes were identified. The genotype frequencies of GH|HaeIII polymorphisms of each breed were shown in Table 1.
In Donggala sheep, AA and AB genotypes had the same genotype frequency of 0.357, and they were higher than BB genotype of 0.286. In East Java sheep AA genotype is the most frequent genotype (0.464), whereas BB and AB were 0.286 and 0.250, respectively.
|
Table 1. Frequency GH|HaeIII genotype in Donggala and East Java sheep |
|||
|
Breed |
Genotype Frequency |
||
|
G1 AA |
G2 BB |
G3 AB |
|
|
Donggala (42)* |
0.357 (15) |
0.286 (12) |
0.357 (15) |
|
East Java (28) |
0.464 (13) |
0.286 (8) |
0.250 (7) |
|
*numbers in the brackets shows sample size |
|||
There is a similarity in pattern of allele frequencies of both breeds, which allele A is higher than allele B (Table 2). The degree of heterozygosity of the GH|HaeIII polymorphisms in Donggala and East Java sheep are 0.503 and 0.498, respectively (Table 2). The degree of heterozygosity represents the mean percentage of heterozygous individuals in a population and describes level of polymorphism of alleles. High heterozygosity shows high genetic diversity within a population (Nei and Kumar 2000).
Based on the heterozygosity values obtained, it shows that the GH|HaeIII polymorphisms had a high degree of genetic diversity. In the selection program, high heterozygosity is desirable, as the high heterozygosity reflects genetic variation of genes in a population and gives a greater opportunity for selection of genes in a population. This suggests that Donggala sheep have greater opportunity for genetic improvement than East Java sheep, if selection program for this marker were implemented.
|
Table 2. Allele frequency and degree of heterozygosity of GH|HaeIII gen in Donggala and East Java sheep |
||||
|
Breed |
Allele Frequency |
Degree of Heterozygosity |
||
|
A |
B |
|
||
|
Donggala (42) |
0.536 |
0.464 |
|
0.503 |
|
East Java (28) |
0.589 |
0.411 |
|
0.493 |
The results of this study showed that GH|HaeIII polymorphism does not affect birth weight both in Donggala and East Java sheep (Table 3). The absence of GH|HaeIII polymorphism effect on birth weight in the present study is parallel to the study of Moradian et al (2013) in the Makooei sheep, using PCR-SSCP method in detecting polymorphism. The lack of GH genotype effect on birth weight in the present study and those of Moradian et al (2013) could be explained that the fetus growth is more under nongenetic maternal environmental influences, rather than the genotype effect of the fetus (Gluckman and Pinal 2003).
GH|HaeIII polymorphism affects growth rate and weaning weight both in Donggala and East Java sheep. In Donggala sheep, AA was observed to have significantly higher growth rate than BB. AB, as a heterozygote, did not show any significant differences in growth rate compared to both homozygotes (AA and BB) (Table 3). In East Java sheep, AB was noticed to have significantly higher and lower growth rates than BB and AA, respectively.
The pattern of relationship between GH|HaeIII polymorphism and weaning weight seems to be similar to those and growth rate. Genotype AA showed significantly higher weaning weight than genotypes BB both in Donggala (11.6 kg vs 9.68 kg) and East Java (10.83 kg vs 9.37 kg) sheep. Genotype AB did not show significant differences in weaning weight compared to genotypes AA and BB both in Donggala and East Java sheep (Table 3).
| Table 3. Average growth traits of Donggala and East Java Sheep according to their GH|HaeIII genotypes | ||||||
| Breed |
Sample size and Growth Traits |
Genotype |
SEM | Prob. | ||
|
G1 AA |
G2 BB |
G3 AB |
||||
|
Donggala |
Birth weight (kg)) |
2.44 |
2.18 |
2.39 |
0.060 |
0.205 |
|
|
Growth rate (g/d) |
102a |
83.3b |
867ab |
3.18 |
0.038 |
|
|
Weaning weight (kg) |
11.6a |
9.68b |
10.22ab |
0.303 |
0.024 |
|
East Java |
Birth weight (kg) |
2.78 |
2.96 |
2.73 |
0.072 |
0.460 |
|
|
Growth rate (g/d) |
89.5a |
71.2b |
66.3b |
3.56 |
0.008 |
|
|
Weaning weight (kg) |
10.8a |
9.37b |
8.72ab |
0.322 |
0.012 |
|
* Growth trait data are not shown because the number of sample is not representative of the genotype Means within the same row with different superscripts are different at p<0.05 |
||||||
From the view point of growth rate and weaning weight, sheep with AA HaeIII genotype is preferred by the farmer to BB HaeIII genotype. The height of allele frequency of AA compared to BB in both breeds (Table 2) might could be explained from this view point that the selection tended to favor the preferred AA HaeIII genotype and to cull BB HaeIII genotype, resulting in an increase in frequency of allele A and a decrease in frequency of allele B.
The presence of GH gene polymorphism effects on growth rate and weaning weight in the current study are in accordance with the results of Tahmoorespur et al (2011) and Moradian et al (2013). Tahmoorespur et al (2011) showed associations between SSCP polymorphisms at the GH gene and breeding value for six month weight in Baluchi sheep. Moradian et al (2013) found five SSCP patterns of GH gene and showed an association between those SSCP patterns with six month weight in Makooei sheep.
GH in affecting growth rate and thus weaning weight is mediated by increased intestinal calcium absorption (Boyd and Bauman 1989), increased protein synthesis and deposition (Hart and Johnson 1986; Gluckman et al 1987) and stimulating chondrocyte proliferation (Boyd and Bauman 1989). The present study comes with a finding that that the GH|HaeIII genotype is polymorphic and associates with growth rate and weaning weight.
The presence of associates of genotype of GH|HaeIII on growth rate and weaning weight and the height of degree of heterozygosity of GH|HaeIII polymorphismsin the present study bring to a close that GH|HaeIII polymorphisms can be a genetic marker in sheep selection program for improvement of growth rate and weaning weight. Genotype AA, bands with a restricted fragment of 277 bp could be the marker for the expected growth rate.
We are grateful to Ministry of National Education of Republic of Indonesia for the financial support. The skilled technical supports of Miss Susiati of Biological Molecular Laboratory of Brawijaya University are appreciated.
Ayuk J and Sheppard M C 2006 Growth hormone and its disorders. Postgraduate Medical Journal 82:24–30. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2563724/pdf/24.pdf
Boyd R D and D E Bauman 1989 Mechanisms of action for somatotropin in growth. In: Campion D R Campion,. Hausman G J and Martin R J Martin (Eds.), Current Concepts of Animal Growth Regulation, Plenum Press, New York.
Statistics Indonesia of the Republic Indonesia 2012. http://www.bps.go.id/tab_sub/view.php?kat=3&tabel=1&daftar=1&id_subyek=24¬ab=12
Eisemann J H, Tyrrell H F, Hammond A C, Reynolds P J, Bauman D E, Haaland G L, McMurtry J P and Varga G A 1986 Effect of Bovine growth hormone administration on metabolism of growing Hereford heifers: dietary digestibility, energy and nitrogen balance. The Journal of Nutrition 116: 157-163. http://jn.nutrition.org/content/116/1/157.full.pdf
Gluckman P D and Pinal C S 2003 Regulation of fetal growth by the somatotrophic axis1. The Journal of Nutrition 133: 1741S–1746S. http://jn.nutrition.org/content/133/5/1741S.full.pdf
Gluckman P D, Breier B H and Davis S R 1987 Physiology of the somatotropic axis with particular reference to the ruminant. Journal of Dairy Science 70: 442-466. http://download.journals.elsevierhealth.com/pdfs/journals/0022-0302/PIIS0022030287800280.pdf?refuid=S0301-6226(04)00006-5&refissn=0301-6226&mis=.pdf
Gul F, Sadaf S and Akhter M W 2009 Ovis aries breed lohi growth hormone gene, complete cds. NCBI number accession GenBank: GQ452268. http://www.ncbi.nlm.nih.gov/nuccore/GQ452268
Hajihosseinlo A, Semsarnejad A, Abollow E, Hashrafi F and Negahdary M 2013 Effect of GH gene polymorphisms on biometric traits in Makooei sheep. Annals of Biological Research 4(6): 351-355. http://scholarsresearchlibrary.com/ABR-vol4-iss6/ABR-2013-4-6-351-355.pdf
Hart I C and Johnson I D 1986 Manipulation of milk yield with growth hormone. In: Haresign W and Cole D J A (Eds), Recent Advances in Animal Nutrition, Butterworths, London, UK.
Husain M H, Rusdi, Y Duma, Hafsah and Mirajuddin 1992 The effects of coat color and length on heat tolerance of Donggala fat tailed sheep. Research Report Tadulako University, Palu, Indonesia.
Husain M H 2001. Current information on Donggala fat tailed sheep of and some efforts to purify their genetic. Paper presented in Communication Forum of Animal Husbandry High Education Leaders, 12-15 November 2001. Denpasar, Bali, Indonesia.
Kuulasma T 2002 Oligo Explorer. University of Kuopio, Kuopio, Finland.
Malewa A 2009 Sifat kualitatif eksternal domba Donggala pada tiga lokasi di Sulawesi Tengah. Agrisains 10(2): 101-109.
Mason I L 1980 Prolific Tropical Sheep. FAO Animal Production and Health Paper 17. Food and Agriculture Organization of the United Nations. Rome. http://www.fao.org/docrep/004/x6517e/X6517E04.htm
Minitab 2003 Minitab user’s guide. Data analysis and quality tools. Release 14 for windows. Minitab Inc, Pennsylvania, USA.
Moradian C, Mohamadi N, Sheshdeh S A R, Hajihosseinlo A and Ashrafi F 2013 Effects of genetic polymorphismat the growth hormone gene on growth traits in Makooei sheep. European Journal of Experimental Biology 3(3): 101-105. http://pelagiaresearchlibrary.com/european-journal-of-experimental-biology/vol3-iss3/EJEB-2013-3-3-101-105.pdf
Neathery M W, Crowe C T, Hartnell G F, Veensuizen J J, Reagan J O and Blackmon D M 1991 Effects of sometribove on performance, carcass composition and chemical blood characteristics of dairy calves. Journal of Dairy Sciece 74: 3933-3939. http://download.journals.elsevierhealth.com/pdfs/journals/0022-0302/PIIS002203029178586X.pdf
Nei M and Kumar S 2000 Molecular Evolution and Phylogenetics. Oxford Univ Press, New York.
Noor R R, Djajanegara A and SCHÜLER L 2001 Selection to improve birth and weaning weight of Javanese fat tailed sheep. Arch. Tierz. Dummerstorf, 44 (6): 649-659. http://arch-anim-breed.fbn-dummerstorf.de/pdf/2001/at01p649.pdf
Steel R G D and Torrie J H 1993 Principles and Procedures of Statistics. Second Edition. McGraw-Hill Book Co. Inc. Singapore.
Tahmoorespur M, Taheri A, Gholami H and Ansary M 2011 PCR-SSCP variation of GH and STAT5A genes and their association with estimated breeding values of growth traits in Baluchi sheep. Animal Biotechnology 22 (1): 37-43.
Tahmoorespur M and Ahmadi H 2012 A neural network model to describe weigh gain of sheep from genes polymorphism, birth weight and birth type. Livestock Science 148: 221-226.
Received 9 April 2014; Accepted 14 May 2014; Published 1 June 2014