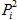 , where
Pi is the frequency of allele i) and Hardy-Weinberg equilibrium were
calculated using PopGene32 program (ver 1.31, Canada).
, where
Pi is the frequency of allele i) and Hardy-Weinberg equilibrium were
calculated using PopGene32 program (ver 1.31, Canada).The following fixed effects model was employed for calculation of BV (breeding value) with DFREML software.
| Livestock Research for Rural Development 24 (9) 2012 | Guide for preparation of papers | LRRD Newsletter | Citation of this paper |
The objective of this study was association of single nucleotide polymorphisms (SNP) in leptin gene with growth traits in Makooei sheep. Genomic DNA was extracted from whole blood samples collected from 130 sheep. PCR products were subjected to SSCP denaturation and polyacrylamide gel electrophoresis. SSCP bands were visualized with silver staining. Five SSCP patterns, representing five different genotypes, were identified.
The evaluation of an association effect between these genotypes with birth weight, weaning weight, 6 month weight, 9 month weight and average daily gain from birth to weaning, weaning to 6 month, 6 to 9 month age, 9 month to yearling weight suggest a positive effect of genotype L4 (BC) with 6 month weight, additive estimated breeding value for the weaning weight, 6 month weight, average daily gain from weaning to 6 month (GWS). In addition, the animal with genotype L4 (BC) had a superior performance for total traits and also the genotype L3 (AC) had a superior birth weight when compared to those of individuals with other pattern. Generally, the BC genotype and B allele were improved growth traits than other genotypes and alleles in Makooei sheep. This result may be related to the leptin large role in post weaning weight that can be affected environmental factors such as nutrition.
Key words: EBVs, PCR-SSCP, polymorphism, weaning weight
The native breeds, because of their natural selection against harsh environment and adaptation to regional conditions are important to resource-poor farmers and pastoralists. Makooei sheep is classified as fat-tailed sheep, similar to Turkish White Karaman, and represented one of the important multi-purpose sheep resources in East and West Azerbaijan provinces of Iran. They are multi colored: black, white with black spots on face and feet. A live female sheep weight about 47 kg. Birth weight of the lambs is 3.7–3.5 (male/females) kg. Indigenous genetic resources of the world are at the risk of extinction due to absorbent crossing with commercial breeds (FAO 2007).The leptin is a hormone synthesized by the adipocyte tissue and regulates the feed intake and the long term regulation of body weight in many species as well as in ruminants. It is highly correlated with body weight and adiposity. The leptin gene is a potential molecular marker because it is related to feed intake, a trait that is difficult to be measured especially in systems based on pastures. The trait also has high economic value and is correlated with production traits. Leptin is a 167-amino acid protein produced by the leptin gene (LEP), plays a key role in regulating energy intake and expenditure, including appetite and metabolism. It is synthesized by adipose tissue and involved in regulation of food intake, energy balance, fertility and immune functions (Fruhbeck et al 1998). The physiological role and biology of leptin is well reviewed (Houseknecht et al 1998; Liefers et al 2003) observed that mutations in the leptin gene or its promoter are associated with differences in serum leptin concentrations and other economically relevant traits in beef and dairy cattle. Liefers et al (2003) also, suggested that dry matter intake, live weight, milk yield, and energy balance affect the serum leptin concentration. Endocrine regulation of leptin, gene expression and secretion has been studied (Macdougald et al 1995). Polymorphisms in the bovine LEP gene have been described (Haegeman et al 2003; Leitman and Read 1997; Pomp et al 1997) and its association with food intake (Lagonigro et al 2003), milk production (Buchanan et al 2003; Liefers et al 2003) carcass and meat quality traits (Schenk el et al 2005) have been reported. However, association studies involving markers in the leptin gene have shown inconclusive results. It could be argued that the inconsistency is due to differences in environmental factors such as nutrition and also to genetic background of the animals, which are potential sources of genetic × environment interactions. Nutritional level, both in terms of quantity and quality of nutrients, becomes an important constraint in production systems that strongly depend on direct grazing (Corva et al 2009). Currently, limited information about exon 3 of leptin gene is available in ovine populations of Iran. Therefore, the objective of this study was to characterize potential variation in the Makooei sheep LEP gene using PCR–single-strand conformational polymorphism (PCR–SSCP) analysis and association with growth traits in Makooei sheep.
Makooei sheep were examined in this study were fat-tailed sheep with medium body size, white color with black spots on face and feet. They are kept in east and west Azerbaijan provinces of Iran and their main products are meat and wool. Blood samples (approximately 2-3 ml) were obtained from 130 unrelated Makooei sheep from different parts of west Azerbaijan province and stored in EDTA-coated tubes. Genomic DNA was extracted from 0.3 ml blood using the genomic DNA purification kit (Cat. No 0512, Fermentas, EU) according to manufacturer's instructions. Quality and quantity of extracted DNA was measured by 0.8 % Agarose gel electrophoresis.
Data were collected at the Makooei Sheep Breeding Station at Makoo city (36o, 35´S and 48o, 22´E) in west-Azerbaijan province. Animals are kept on natural pasture during spring, summer and autumn seasons. Range conditions are poor during the winter months and, therefore, animals are kept indoors during the winter. In general, the flock is managed under a semi-migratory system.
In this study was used to exon 3 of leptin gene due to the ability to identify polymorphism in this area of leptin gene. In addition to, the previous research in other breeds of Iranian sheep was done on this area of leptin gene, that can help in the comparison. The DNA amplification of the LEP gene was achieved by PCR. Two PCR primers, LEP-up (5-AGGAAGCACCTCTACGCTC-3) and LEP-dn (5´-CTTCAAGGCTTCAGCACC-3´), targeting a fragment of 471 bp was employed as described (Zhou et al 2009). The PCRs were carried out in 50 µl volumes using PCR master mix kit (Cinnagen, Iran) containing 2.5 units Taq DNA Polymerase in reaction buffer, 4mM MgCl2, 50 µM each of dATP, dCTP, dGTP and dTTP, 0.5 µM of each primer and about 100 ng of extracted DNA as template. The thermal profile consisted of 2 min at 94oC, followed by 35 cycles of 30 s at 94oC, 30 s at 59oC and 30 s at 72oC, with a final extension of 5 min at 72oC. Amplification was carried out in Master cycler (Eppendorf, Germany).
PCR products were mixed with 8 µl of denaturing loading dye [95% (w/v) deionized formamide, 0.05% (w/v) xylene cyanol, 0.05% (w/v) bromophenol blue and 0.02 M EDTA] in a total volume of 15 µl. The mixture was denatured at 95oC for 5 minutes and was snap chilled on ice. The total volume was applied in a 15% polyacrylamide gel, as described by Herring et al 1982. The electrophoresis was performed in 0.5 X TBE buffer (Tris 100 mM, Boric Acid 9 mM, EDTA 1 mM) at room temperature (18oC) and constant 200 V for 3 h. Polyacrylamide gels were stained with silver according to the protocol described (Herring et al 1982).
The allelic and genotypic frequencies, expected means, observed and expected
Nei’sheterozygosities,
(HE=1-Σ, where
Pi is the frequency of allele i) and Hardy-Weinberg equilibrium were
calculated using PopGene32 program (ver 1.31, Canada).
The following fixed effects model was employed for calculation of BV (breeding
value) with DFREML software.

Where: Yijklmno= dependent variable evaluated on the ith level of the
random factor; year (YRi , i=1, 2, 3,…, 21), the jth level of the
fixed factor; sex (SXj , j=1 and 2), the kth level of the fixed
factor; number of offspring in each birth (BTk , K= 1,2 and 3), the
lth level of the fixed factor; mother age (ADl , l=1,2,…,7), mth
level of the random additive genetic effect (ANm, m= number of animal
for each trait), µ = overall mean for each trait, PEn, random effect
of maternal permanent environment in n level and eijklmno is the
random error effect. The estimated parameters according to model were:
phenotypic variance(σ2p ), direct additive genetic
variance(σ2a ), residual variance (σ2e
), direct heritability(h2a), (σ2a/σ2p).
For the association studies, the traits of interest were analyzed using the
general linear model (GLM) procedure of the SAS program, according to the
following statistical model:
Yijk= μ+ Gi+Sj+β(BWij-(BW) +eijk
Yijkl= production trait, μ = the overall mean, Gi= the fixed effect
of the ith genotype for leptin (i= 1, 2,…,5), Sj= the fixed effect of
sex (j = 1, 2), β=birth weight effect as linear covariable for all variables
except BW, eijkl = the random residual error.
All extracted DNAs from sheep blood samples yielded a specific single band PCR product without any nonspecific band. The PCR-SSCP analysis of exon3 leptin revealed five distinct patterns. The frequencies of the observed genotypes were 0.0 9, 0.17, 0.37, 0.14 and 0.23 for BB, AB, BC, AC and CC, respectively. Allele frequencies were 0.15, 0.37 and 0.48 for A, B and C respectively.
The expected homozigosity (Homexp), expected heterozygosity (Hexp), observed heterozygosity (Hobs) , observed homozigosity (Obs-Hom) and average heterozygosityvalues for leptinwere0.3845, 0.6155, 0.6769, 0.323, 0.6108, respectively. The chi-square test showed no significant deviation (P>0.05) from Hardy-Weinberg equilibrium for this locus in studied population.
The evaluation of an association effect between these SSCP patterns with birth weight (BW), weaning weight (WW), six month weight (SW), nine month weight (NW), and average daily gain from birth to weaning (GBW), weaning to six month (GWS) , from six month to nine month age (GSN), from nine month to yearling weight (GNY) suggest a positive effect of genotype L4 (BC) with 6 month weight, additive estimated breeding value for the weaning weight, 6 month weight, average daily gain from weaning to 6 month (GWS). In addition, the sheep with genotype L4 (BC) had a superior performance for WW, SW NW and YW and also the genotype L3 (AC) had a superior birth weight when compared to those of individuals with other pattern (Table1).
The estimated effects suggest that most of the impact of LEP is realized after weaning. Leptin regulates appetite and energy metabolism by decreasing food intake and increasing energy expenditure and physical activity (Houseknecht et al 1998). Functionally, leptin is best known for its role in regulating rodent appetite and energy expenditure. Leptin has been associated with fat deposition in cattle (Buchanan et al 2003). In cattle a single nucleotide polymorphism (SNP) i.e. a cytosine to thymine transition results in an amino acid change from arginine to cysteine was responsible for the increased carcass fat deposition (Buchanan et al 2003). In sheep, leptin blood levels are associated with feeding levels (Marie et al 2001),diet nutritional value (Blache et al 2000) and body fat mass (Delavaud et al 2002).Previous studies have shown associations of LEP gene polymorphisms and leptin blood levels with carcass and meat quality traits in beef cattle (Buchanan et al 2003). There are few articles on exon 3 polymorphism of leptin gene in sheep. The polymorphism in the exon 3 of the LEP in sheep was also reported by Zhou et al (2009) using PCR-SSCP technique. They reported five unique SSCP patterns corresponding to five allelic sequences using the combination of two different electrophoresis conditions in six common New Zealand sheep breeds. In another study leptin gene polymorphism and its association with skeletal muscle growth and meat quality was investigated using single nucleotide polymorphism (SNP) analysis. A total of three SNPs were identified in the ovine LEP gene including two SNP in intron 2 and one in the 3’UTR (Boucher et al 2006).
There are several studies on the association of LEP gene polymorphism and other traits in animals. Almeida et al (2003) reported that RFLP genetic marker genotypes in the leptin gene were significantly associated with calving date in a Brangus herd in Brazil. A weak correlation between leptin serum levels and cow body condition score (BCS) and a strong relationship between leptin serum levels and the size of adiposities has been described (Delavaud et al 2002). Madeja et al (2004) reported that the HphI polymorphism has a significant effect on milk and protein yield. Animals with the TT genotype had approximately 2×higher estimated breeding values for milk and protein yields. Nkrumah et al (2004) reported an association of the T allele with higher carcass grades but lower carcass lean percentage, with no effect on feed intake or feed efficiency. Most association studies involving leptin have focused on carcass composition and beef quality traits. Di Stasio et al (2007) reported one of the few studies that included growth traits. In a sample of 59 Blonde d’Aquitaine bulls, the C allele at SNP2 had a favorable effect on weight gain. Very little information is currently available to compare different ovine populations from Iran. In this study, variation in the exon 3 coding sequence of the ovine LEP gene was investigated using PCR–SSCP analysis. In the present study three alleles (A, B, and C) and five genotypes (AC, AB, BB, BC, and CC) were identified for exon III of LEP gene in "Makooei" sheep in west Azerbaijan, Iran. The most frequent allele and genotype in the "Makooei" sheep were allele C and genotype BC with frequencies of 48% and 37%, respectively.
The studies about leptin (exon 3) genotypes were carried out by Tahmoorespur et al (2009) and Shojaei et al (2010) in Baluchi and kermani sheeps of Iran, respectively. They analyzed the same exon with the same methodology and primers and found 3 genotypes in the Baluchi and kermani sheeps, but, in this study detected five genotypes for exon 3 of Leptin gene according to Zhou et al (2009). Variations detected here might have an impact on leptin activity and function. In the tested Makooei sheep population, significant statistical results were found in SW (six month weight), additive estimated breeding value for the weaning weight (WW), six month weight (SW) and this result may be related to the leptin large role in post weaning weight that may affected environmental factors such as nutrition. These results also show that the leptin gene is involved in the long-term regulation of body weight. Only a few studies have been conducted to investigate the association between leptin gene polymorphism and performance traits in sheep. Tahmoorespur et al (2009) analyzed the association of leptin polymorphism with average daily gain (ADG) of Baluchi sheep. They reported a significant association between leptin polymorphism and ADG at birth to 3 months of age (P<0.10). The association of leptin polymorphism with the growth traits in Kermani sheep (Shojaei et al 2010) indicates that the growth traits are significantly affected by the genotypes.
|
Table 1: Least square means of the breeding values, growth traits and daily gain of Makooei sheep according to the SNP genotype in Leptin |
|||||||||
|
Genotypes |
|||||||||
|
|
L1(AA) |
L2(AB) |
L3(AC) |
L4(BC) |
L5(CC) |
SEM |
Prob. |
||
|
Breeding values (Kg) |
|
|
|||||||
|
BW |
-0.005 |
0.035 |
-0.013 |
0.042 |
0.085 |
0.040 |
0.105 |
||
|
WW |
1.56ab |
1.52ab |
0.760 a |
2.12 b |
1.72ab |
0.325 |
0.033 |
||
|
SW |
1.39ab |
2.03ab |
0.470 a |
2.46 b |
1.30ab |
0.664 |
0.024 |
||
|
NW |
0.783 |
0.927 |
0.195 |
0.965 |
0.544 |
0.292 |
0.073 |
||
|
YW |
0.783 |
0.927 |
0.195 |
0.965 |
0.544 |
0.292 |
0.073 |
||
|
Growth traits (Kg) |
|
|
|||||||
|
BW |
3.67 |
3.83 |
4.06 |
3.85 |
3.93 |
0. 124 |
0.470 |
||
|
WW |
19.9 |
19.6 |
20.0 |
20.9 |
19.7 |
0.622 |
0.230 |
||
|
SW |
28.5 a |
28.9ab |
29.0ab |
31.3 a |
23.0 b |
1.01 |
0.0040 |
||
|
NW |
29.4 |
28.7 |
28.2 |
29.7 |
29.0 |
1.48 |
0.461 |
||
|
YW |
35.8 |
34.9 |
36.9 |
38.6 |
37.8 |
1.54 |
0.170 |
||
|
Daily gain (kg/day) |
|
|
|||||||
|
GBW |
0.174 |
0.171 |
0.175 |
0.186 |
0.171 |
0.007 |
0.350 |
||
|
GWS |
0.094ab |
0.103ab |
0.099ab |
0.116a |
0.094b |
0.008 |
0.032 |
||
|
GSN |
0.010 |
-0.004 |
-0.008 |
-0.017 |
0.008 |
0.013 |
0.143 |
||
|
GNY |
0.084 |
0.063 |
0.081 |
0.097 |
0.095 |
0.011 |
0.094 |
||
|
Birth weight (BW), weaning weight (WW), six month weight (SW), nine month weight (NW), yearling weight (YW), average daily gain from birth to weaning (GBW), weaning to six month (GWS) , from six month to nine month age (GSN) and from nine month to yearling weight (GNY) abMeans in the same row without common letter for each effect are significant different . |
|||||||||
According to table 2, the high genetic and phenotypic correlation between
weaning weight with 9 month weight and yearling weight indicates that a breeding
rams may be selected at an earlier age for the LEP gene. Genetic correlations
between the direct effects on all traits were positive and ranged from 0.32 to
0.93. The results of the present investigation showed that due to positive
genetic and phenotypic correlations, selection for any of the studied traits can
result in an increase of phenotypic magnitudes and genetic potentials for body
weight traits.
|
Table 2: Correlation between growth traits in Iranian 'Makooei' sheep breed |
|||
|
Trait 1 |
Trait 2 |
rp12 |
ra12 |
|
WW |
SW |
0.77 |
0.75 |
|
WW |
NW |
0.62 |
0.56 |
|
WW |
YW |
0.54 |
0.32 |
|
SW |
NW |
0.76 |
0.93 |
|
SW |
YW |
0.59 |
0.81 |
|
NW |
YW |
0.73 |
0.87 |
|
Birth weight (BW), weaning weight (WW), six month weight (SW), nine month weight (NW), yearling weight (YW), rp12: phenotypic correlation between trait1 and trait2; ra12: direct additive genetic correlation between trait 1 and trait 2. |
|||
Results of this study showed that partly are in accordance with the results of Tahmoorespur et al (2010) that effect of the leptin gene was significant with weaning weight (WW). To date, this was the (secondly) study that attempted to detect allele variation in the ovine LEP gene and its association with growth traits in Iranian sheep breeds. Further studies will be needed before application of this SNP to the breeding industry. More population studies and functional analyses of these genetic variations will be needed to reveal the genotype-phenotype relationships in sheep. It is advisable to apply more markers, in addition to the applied markers, in this breed and some other native breeds such as Ghezel and Haraky in order to uncover their genetic relationship.
We thank the Makooei Sheep Breeding Station (MSBS) staff for providing us the data used in this study. Also we are grateful to the head of the institute of biotechnology, Urmia University for providing laboratory facilities.
Almeida S E, Almeida E A, Moraes J C F and Weimer T 2003 Molecular marker in the LEP gene and reproductive performance of beef cattle. Journal of Animal Breeding and Genetic 120: 106-113. http://onlinelibrary.wiley.com/doi/10.1046/j.1439-0388.2003.00377.x/full
Blache D, Tellam R L, Chagas L M, Blackberry M A, Vercoe P E and Martin G B 2000 Level of nutrition affects leptin concentrations in plasma and cerebrospinaluid in sheep. Journal of Endocrinology 165:625-637.
Boucher D, Palin M F, Castonguay F, Gariépy C and Pothier F 2006 Detection of polymorphisms in the ovine leptin LEP gene: Association of a single nucleotide polymorphism with muscle growth and meat quality traits. Canadian Journal of Animal Science 86:31-35. http://pubs.aic.ca/doi/pdf/10.4141/A05-052
Buchanan F C‚ Van Kessel A G‚ Waldner C‚ Christensen D A‚ Laarveld B and Schmutz S M 2003 An association between a leptin single nucleotide polymorphism and milk and protein yield. Journal of Dairy Science 86: 3164-3166. http://download.journals.elsevierhealth.com/pdfs/journals/0022-0302/PIIS0022030203739186.pdf
Corva P M, Fernández Macedo G V, Soria L A, Papaleo Mazzucco J, Motter M, Villarreal E L, Schor A, Mezzadra C A, Melucci L M and Miquel M C. 2009 Effect of leptin gene polymorphisms on growth, slaughter and meat quality traits of grazing Brangus steers. Genetics and Molecular Research 81: 105-116. http://www.ncbi.nlm.nih.gov/pubmed/19283678
Delavaud C, Ferlay A, Faulconnier Y, Bocquier F, Kann G and Chilliard Y 2002 Plasma leptin concentration in adult cattle: Effects of breed, adiposity, feeding level, and meal intake. Journal of Animal Science 80: 1317-1328. http://jas.fass.org/content/80/5/1317.full.pdf
Di Stasio L, Brugiapaglia A, Galloni M and Destefanis G and Lisa C 2007 Effect of the leptin c.73T>C mutation on carcass traits in beef cattle. Animal Genetics 38: 316-317. http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2052.2007.01595.x/abstract
FAO 2007
Report on the International
Technical Conference on Animal Genetic Resources for Food and Agriculture,
Interlaken Switzerland, September 2007.
http://www.fao.org/ag/againfo/programmes/en/genetics/angrvent2007.html
Fruhbeck G, Jebb S A and Prentice A M 1998
Leptin: physiology and pathophysiology. Journal of Basic and Clinical Physiology
and physiology 18: 399-419.
http://onlinelibrary.wiley.com/doi/10.1046/j.1365-2281.1998.00129.x/abstract
Haegeman A, Van Zeveren A and Peelman L J 2003 New mutation in exon 2 of the bovine leptin gene. Animal Genetics. 31- 79.
Herring A J, Inglis N F, Ojeh C K, Snodgrass D R and Menzies J D 1982 Rapid diagnosis of rotavirus infection by direct detection of viral nucleic acid in silver-stained polyacrylamide gels. Journal of Clinical Microbiology. 16: 473-477. http://jcm.asm.org/content/16/3/473.full.pdf
Houseknecht K L, Baile C A, Matteri R L, Spurlock M E 1998 The biology of leptin. Journal of Animal Science 76: 1405-1420. http://jas.fass.org/content/76/5/1405.full.pdf
Lagonigro R, Wiener P, Pilla F, Woolliams J A and William J 2003 A new mutation in the coding region of the bovine leptin gene associated with feed intake. Animal Genetics 34(5): 371-374. http://onlinelibrary.wiley.com/doi/10.1046/j.1365-2052.2003.01028.x/full
Leitman S F and Read E J 1997 Hematopoietic progenitor cells. Seminars Hematology. 83: 341-358. http://serials.unibo.it/cgi-ser/start/it/spogli/df-s.tcl?prog_art=2190367&language=ITALIANO&view=articoli
Liefers S C, Te Pas M F, Veerkamp R F, Chilliard Y, Delavaud C, Gerritsen R and Van Der Lende T 2003 Association of leptin gene polymorphisms with serum leptin concentration in dairy cows. Mammalian Genome 14: 657–663. http://www.ncbi.nlm.nih.gov/pubmed/14629116
Macdougald O A, Hwang C, Fan H and Lane M D 1995 Regulated expression of the obese gene productleptin) in white adipose tissue and 3T3-L1 adipocytes. Proceedings of the National Academy of Sciences 92: 9034-9037. http://www.ncbi.nlm.nih.gov/pmc/articles/PMC40918/pdf/pnas01498-0032.pdf
Madeja Z, Adamowicz T, Chmurzynska A, Jankowski T, Melonek J, Switonski M and Strabel T 2004 Effect of leptin gene polymorphisms on breeding value for milk production traits. Journal of Animal Science 87:3925–3927. http://jds.fass.org/cgi/reprint/87/11/3925.pdf
Marie M, Findlay P A, Thomas L and Adam C L 2001 Daily patterns of plasma leptin in sheep: effects of photoperiod and food intake. Journal of Endocrinology 170, 277-286. http://joe.endocrinology-journals.org/content/170/1/277.full.pdf
Nkrumah J D, Li C, Basarab J B, Guercio S, Meng Y, Murdoch B, Hansen C and Moore S S 2004 Association of a single nucleotide polymorphism in the bovine leptin gene with feed intake, feed efficiency, growth, feeding behaviour, carcass quality and body composition. Canadian Journal of Animal Science 84: 211-219. http://pubs.aic.ca/doi/pdf/10.4141/A03-033
Pomp D, Zou T, Clutter A C and Barendse W 1997 Mapping of leptin to bovine chromosome 4 by linkage analysis of a PCR based polymorphism. Journal of Animal Science 75:1427. http://www.animal-science.org/content/75/5/1427.full.pdf
Schenkel F S, Miller S P, Ye X, Moore S S, Nkrumah J D, Li C, Yu J, Mandell I B, Wilton J W and Williams J L 2005 Association of single nucleotide polymorphisms in the leptin gene with carcass and meat quality traits of beef cattle. Journal of Animal Science 83: 2009-2020. http://www.ncbi.nlm.nih.gov/pubmed/16100055
Shojaei M, Mohammadabadi M R, Asadi Fozi M, Esmailizadeh K A, Ferdowsi M H, Torabi A, Tayyarzadeh M and Mirzakhani H 2010 Using PCR-SSCP technique to investigate polymorphism of Leptin gene in Kermani sheep. Animal Science Researches 20: 115-122. (UJ3khC1hzQEkAAAAZTU4MDBlYmMtZWYzZS00NWI3LWIzM2YtYTg4ODU0MTU5NTk3_nWtuX1Mz_hu3t6NQjlR6g5UxkQ1))/Uploads/Journal/PublishedArticles/90F3_41.pdf
Tahmoorespur M, Nassiry M R, Ansary M, Heravi Mousavi A, Vafaye vale M and Eftekhari Shahroudi F 2009 Analysis of Leptin gene polymorphism and their association with average daily gain trait in Baluchi sheep. 3rd Congress of animal science, Mashhad, Iran.
Tahmoorespur M, Taheri A, Vafaye Valeh M, Nassiry M R and Ansary M 2010 Assessment relationship between leptin and Ghrelin genes polymorphisms and estimated breeding values EBVs) of growth traits in Baluchi sheep. Journal of Animal and veterinary Advances 9(19): 2460-2465. http://docsdrive.com/pdfs/medwelljournals/javaa/2010/2460-2465.pdf
Zhou H, Hickford H and Gong H 2009 Identification of allelic polymorphism in the ovine leptin gene. Molecular Biotechnology 41: 22–25. http://rd.springer.com/article/10.1007/s12033-008-9090-3
Received 7 August 2012; Accepted 16 August 2012; Published 3 September 2012