|
|
|
|
| Livestock Research for Rural Development 21 (9) 2009 | Guide for preparation of papers | LRRD News | Citation of this paper |
Four Lai Sind bulls (26-28 months of age and 290 kg average live weight with permanent rumen cannulas were fed a basal diet of natural grass and levels of dried cassava root peelings (RP) of: 0, 0.25, 0.50 and 0.75 kg DM per 100 kg LW. These treatments were applied according to a 4*4 Latin Square arrangement with periods of 30 days on each treatment.
Increasing the offer of RP resulted in: reduced intake of the grass but increased overall intake of DM; a reduced concentration of rumen ammonia with minimal effects on rumen pH; an increase in the rumen protozoal population but a reduction in that of bacteria; a decrease in the rumen degradability of grass DM with minimal effects on the degradability of the cassava root peelings.
Despite the apparently negative effects on the balance of rumen bacteria and protozoa, the overall impact of the RP supplementation appeared to be a better balance of nutrients to the animal as reflected in the linear increase in total DM intake.
Key words: ammonia, bacteria, cannula, degradability, in sacco, pH, protozoa
A siginificant amount of dried cassava leaves (0.96 tonnes/ha) and peelings (1.75 tons/ha) are obtained at root harvesting, but these are underutilized and usually left on the soil for decomposition (Ravindran and Rajaguru 1988; Dương Nguyen Khang and Wiktorsson 2000). A limiting factor for their utilization as feedstuffs may be the high content of cyanogenic glucosides, which produce the cyanide (HCN) toxin. Cyanide concentrations of nearly 1,000 mg kg-1 dry matter (DM) have been reported in cassava forage (Man and Wiktorsson 2001). Earlier reports indicated that feeding large amounts of untreated cassava products could result in the death of animals, particularly non-ruminants (Hill 1973). In cattle and sheep, HCN can be lethal at levels of 2-4 mg HCN kg-1 body weight (Kumar 1992). However, rumen bacteria have the capacity to hydrolyze β-D-glucosides (Majak and Cheng 1984; Majak et al 1990) and to utilize the glucose as an energy source. The toxic HCN can be rapidly absorbed, eructated or further metabolized in the rumen.
Sun-drying alone eliminated almost 86% of the initial HCN content in cassava leaves (Devendra 1977; Gomez and Valdivieso1988). When combining chopping and wilting, HCN in the dried cassava leaves meal was reduced to a level which was considered to be safe for monogastric animals (Ravindran et al 1987). Nguyen Thi Thu Hong et al (2003), Doan Duc Vu (2004) and Duong Nguyen Khang (2004) studied dried cassava foliage as a protein source for beef and dairy rations and obtained good results. In practice, farmers have used for long time low levels of dried and wilted cassava peelings for beef cattle feeding, but no formal research on the topic was made. The present study aimed at examining the effects of different levels of dried cassava root peelings (RP) on rumen pH, rumen ammonia and the population of rumen micro-organisms in cattle fed natural grasses as a basal diet.
The experiment was carried out at Can Tho University Experimental Farm, located in Can Tho city, Vietnam. Four Lai Sind bulls (26-28 months of age and 290 kg body weight (BW) on average) with permanent rumen cannulas were used for the experiment. Each animal was individually placed in a pen. Fresh water was available ad libitum during the whole experiment.
The treatments were levels of dried cassava root peelings (RP) of 0, 0.25, 0.50 or 0.75 kg DM/100 kg LW/day (designated as RP0, RP0.25, RP0.5 and RP0.75). The experiment was arranged as a 4*4 Latin Square; each treatment period lasted for 30 days. The first two weeks of each period were for animals and micro-flora adaptation to the new diets. Data on daily feed intake were taken during 7 days of the third week (days 15 to 21). Feed samples for analysis were taken before feeding during the last 3 days of the same week (days 19 to 21); in sacco degradation of feed samples was determined during a 72 h period (days 22 to 24); rumen samples for pH and concentration of ammonia nitrogen were taken during days 28 and 29. The RP were fed once daily according to the treatments throughout the experimental period.
The basal diet consisted of natural grasses offered ad libitum and supplied in two meals daily at 7:00h and 15:00h. Supplements of 1 kg cassava root meal, and 20 g of a mixture of salt and minerals were given once daily at about 7:30h in addition to dried cassava root peelings as mentioned before.
The natural grasses were harvested in the immediate vicinity of the cattle pens two times/day. Cassava root meal was bought on the market at one occasion. The RP was collected at the same time from one field after harvesting the roots. The peels were air-dried on the ground.
Refused feeds were weighed each morning during 7 days of the third week. The feeds were also sampled daily at these occasions for later analysis.
Feed samples were analyzed for crude protein (CP), ether extract (EE), crude fiber (CF), total ash and HCN (RP only) according to the procedure of the AOAC (1990). For the determination of degradable dry matter (DDM) and degradable organic matter (DOM) samples of the grasses and of RP were suspended in the rumen of fistulated animals for up to 72 hrs using the in sacco degradation procedure (Ørskov et al 1980). The bags were 60x120mm and made of nylon filter cloth with a pore size of 28 microns. The bags were attached to plastic tubes and incubated in the rumen for 6, 12, 24, 48 and 72 hours. After incubation, the bags were washed by hand under running tap water until the water ran clear, and then dried in a microwave oven to a constant weight.
On days 28 and 29 of each period, ruminal fluid samples were collected before feeding in the morning and then at an interval of 2 hours over an 8-hour period through a probe placed in a caudal position in the ventral part of the rumen. Protozoa population in the rumen fluid was estimated by diluting at a ratio of ¼ with methylene blue and protozoa were counted under light-microscopy (10 x magnification) using a 0.2 mm deep dollfus counting Mallaze chamber. All fields in the counting chamber were filled for protozoa counting by the Dehority method (1984).
For counting of bacteria, rumen fluid was diluted at the ratio of ¼ with 0.05% formol to make solution A. This solution was then diluted at the ratio of 1:7 with distilled water and centrifuged at 1,500 rpm for 10 minutes to make solution B. One ml of the B solution was diluted with 1 ml of glugol solution and the counting was made using a Neubauer chamber. Five fields in the counting chamber were filled for counting bacteria by the Warner method (1962).
pH and ammonia nitrogen concentration in rumen fluid were determined immediately after collection using a pH meter. The concentration of ammonia nitrogen in the rumen fluid (NH3-N) was determined by distillation and absorption in boric acid followed by titration with H2SO4 0.1N (Preston 1995)
Data were analyzed by ANOVA using the General Linear Models procedure and Pairwise comparisons in Minitab Statistical Software version 12.21 (Minitab 2000). Sources of variation were: treatments, periods, animals and error.
The chemical composition of grass, cassava root meal and RP is shown in table 1. During the experiment period (120 days), there were no changes in the chemical composition of feeds.
|
Table 1. Chemical composition of feed ingredient used in experiment (mean and SE on DM basic) |
|||||||
|
|
Samples |
DM, % |
Nutrient value, %/DM |
HCN, mg/kg DM |
|||
|
CP |
CF |
EE |
Ash |
||||
|
Grass |
21.4±1.38 |
12,20 ±2.64 |
37.00 ±2.87 |
3,00 ±2.84 |
12.6 ±2.2 |
- |
|
|
RP |
93.3±0.93 |
8,20 ±1.95 |
10.00 ±1.25 |
1,10 ±0.16 |
6.24 ±0.54 |
366 ±64.2 |
|
|
Cassava root meal |
4 |
89.1±1.01 |
2.70 ±1.51 |
2,23 ±1.03 |
2.00 ± 0.65 |
2.74 ±0.16 |
4.93 ±0.02 |
Grass intake was decreased but total DM intake was increased (Table 2 and Figure 1) according to the level of dietary RP.
|
Table 2. Mean values for intakes of grass and cassava root peelings (RP) by Lai Sind bulls (DM/day) |
||||||
|
|
RP0 |
RP0.25 |
RP0.5 |
RP0.75 |
SEM |
P |
|
Grass, kg |
6.05a |
5.76ab |
5.50ab |
4.85b |
0.17 |
0.028 |
|
Cassava root meal, kg |
0.89 |
0.89 |
0.89 |
0.89 |
- |
- |
|
RP, kg |
0a |
0.60b |
1.19c |
1.80d |
0.06 |
0.001 |
|
Total DM, kg |
6.94 |
7.24 |
7.59 |
7.53 |
0.16 |
0.182 |
|
DMI, kg/100 kg LW/d |
2.44 |
2.56 |
2.64 |
2.65 |
0.08 |
0.271 |
|
Grass DMI, kg/100 kg LW/d |
2.12a |
2.04ab |
1.91ab |
1.71b |
0.08 |
0.036 |
|
RP as % of total DMI |
0 |
8 |
16 |
24 |
- |
- |
|
HCN intake, mg/kg LW/d |
0 |
0.7 |
1.4 |
2.1 |
- |
- |
|
CP in DM, % |
10.9 |
10.7 |
10.5 |
10.1 |
- |
- |
|
abcd Means in the same row without common superscript are different at P<0.05 |
||||||
|
|
|
|
The ruminal pH was not affected by the feeding of cassava RP at all sampling times except for the 8 h sample when there was an indication (P=0.06) that it increased linearly with increasing dietary RP (Table 3).
|
Table 3. Levels of ruminal pH and NH3-N, number of protozoa and bacterial populations in rumen fluid of Lai Sind bull after feeding |
||||||
|
Hours after feeding |
RP0 |
RP0.25 |
RP0.5 |
RP0.75 |
SEM |
P |
|
pH |
|
|
|
|
|
|
|
0 |
6.39 |
6.34 |
6.27 |
6.70 |
0.070 |
0.122 |
|
2 |
6.36 |
6.30 |
6.33 |
6.69 |
0.080 |
0.274 |
|
4 |
6.32 |
6.29 |
6.32 |
6.55 |
0.080 |
0.760 |
|
6 |
6.23 |
6.19 |
6.26 |
6.38 |
0.080 |
0.926 |
|
8 |
6.15 |
6.15 |
6.22 |
6.30 |
0.070 |
0.907 |
|
NH3-N, mg/litre |
|
|
|
|||
|
0 |
149 |
159 |
159 |
128 |
5.0 |
0.117 |
|
2 |
190 |
178 |
177 |
167 |
6.0 |
0.228 |
|
4 |
172 |
163 |
143 |
134 |
7.0 |
0.250 |
|
6 |
150 |
151 |
134 |
114 |
7.0 |
0.292 |
|
8 |
144 |
145 |
143 |
138 |
10.0 |
0.997 |
|
Bacteria, x10-8/ml |
|
|
|
|||
|
0 |
1.99 |
1.81 |
1.57 |
1.58 |
0.100 |
0.480 |
|
2 |
1.97 |
1.78 |
1.70 |
1.55 |
0.080 |
0.483 |
|
4 |
1.87 |
1.73 |
1.68 |
1.44 |
0.070 |
0.432 |
|
6 |
1.81 |
1.49 |
1.43 |
1.40 |
0.080 |
0.171 |
|
8 |
1.59 |
1.45 |
1.38 |
1.24 |
0.080 |
0.162 |
|
Protozoa, x10-5/ml |
|
|
|
|||
|
0 |
1.59 |
3.08 |
3.56 |
3.29 |
0.420 |
0.060 |
|
2 |
1.38 |
2.52 |
3.10 |
3.10 |
0.310 |
0.069 |
|
4 |
1.69 |
2.65 |
3.70 |
2.72 |
0.300 |
0.280 |
|
6 |
1.05 |
2.13 |
2.52 |
2.16 |
0.250 |
0.122 |
|
8 |
1.00 |
1.70 |
2.49 |
2.10 |
0.310 |
0.144 |
Rumen ammonia concentrations decreased linearly with increasing RP in the diet (Tables 3 and 4; Figure 2).
|
Table 4. Relationship between RP level (kg DM/kg LW) and ammonia concentration in rumen fluid |
|||
|
Time, h |
Equation |
R2 |
P |
|
0 |
NH3-N = 158 – 25.5 RP |
0.31 |
0.443 |
|
2 |
NH3-N = 188 – 28.0 RP |
0.92 |
0.040 |
|
4 |
NH3-N = 173 – 53.6 RP |
0.97 |
0.013 |
|
6 |
NH3-N = 156 – 50.0 RP |
0.87 |
0.070 |
|
8 |
NH3-N = 145 – 8.0 RP |
0.69 |
0.017 |
|
Table 5. Relationship between RP level (kg DM/kg LW) and bacteria numbers (x10-8/ml) in rumen fluid |
|||
|
Time, h |
Equation |
R2 |
P |
|
0 |
Bacteria= 1.96 - 0.59 RP |
0.89 |
0.058 |
|
2 |
Bacteria= 1.95 - 0.54 RP |
0.98 |
0.011 |
|
4 |
Bacteria= 1.88 – 0.54 RP |
0.93 |
0.034 |
|
6 |
Bacteria= 1.73 – 0.52 RP |
0.78 |
0.118 |
|
8 |
Bacteria= 1.58 - 0.45 RP |
0.99 |
0.008 |
|
|
|
|
The values were within the range (150 to 200 mg/litre) considered to be optimal for rumen microbial activity on forage=based diets (Preston and Leng 1987). Decreasing levels of ammonia with feeding of RP indicates a stimulation of rumen microbial protein synthesis in response to the fermentable carbohydrate present in the cassava RP. Cassava root meal has been shown to ferment more rapidly in the rumen than other starch-rich feeds such as maize and rice grains (Fernandez and Hovell 1978).
Protozoal and bacterial populations in the rumen were influenced by the dietary level of RP (Tables 3 and 6; Figures 3 and 4).
|
|
|
Figure 3. Relationship between RP level (kg DM/kg LW) and bacteria numbers (x108/ml) in rumen fluid |
|
|
|
Figure 4. Relationship between diet RP level (kg DM/kg LW) and protozoa numbers (x10-5/ml) in rumen fluid |
Populations of bacteria decreased with increasing level of dietary RP (Table 5) while there were tendencies for protozoal populations to increase (Table 6).
|
Table 6. Relationship between RP level (kg DM/kg LW) and protozoa numbers (x10-5/ml) in rumen fluid |
|||
|
Time, h |
Equation |
R2 |
P |
|
0 |
Protozoa= 2.04 + 2.23 RP |
0.67 |
0.183 |
|
2 |
Protozoa= 1.66 + 2.30 RP |
0.84 |
0.086 |
|
4 |
Protozoa= 2.07 + 1.66 RP |
0.42 |
0.349 |
|
6 |
Protozoa= 1.41 + 1.49 RP |
0.57 |
0.244 |
|
8 |
Protozoa= 1.21 + 1.64 RP |
0.69 |
0.170 |
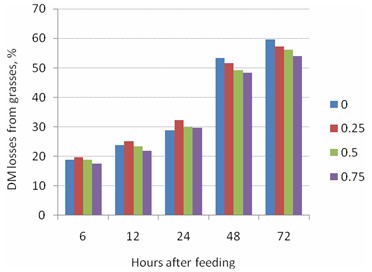
Except for the samples taken after 6h, increasing the level of RP in the diet led to linear decreases in the DM degradability (Tables 7 and 8; Figure 5).
|
Table 7. Mean values for degradable dry matter (DDM) of feed samples after incubation |
||||||
|
After feeding, h |
RP0 |
RP0.25 |
RP0.5 |
RP0.75 |
SEM |
P |
|
|
Grass |
|
|
|||
|
6 |
18.9 |
19.8 |
18.9 |
17.6 |
0.9 |
0.120 |
|
12 |
23.9 |
25.1 |
23.3 |
21.9 |
1.3 |
0.271 |
|
24 |
28.8 |
32.2 |
29.8 |
29.6 |
1.5 |
0.587 |
|
48 |
53.3 |
51.6 |
49.2 |
48.3 |
2.3 |
0.506 |
|
72 |
59.6 |
57.3 |
56.1 |
54.0 |
2.4 |
0.230 |
|
|
Cassava RP |
|
|
|||
|
6 |
15.1 |
17.1 |
20.4 |
17.5 |
2.2 |
0.067 |
|
12 |
23.2 |
22.1 |
21.6 |
19.5 |
1.5 |
0.293 |
|
24 |
26.9 |
29.0 |
28.5 |
25.7 |
1.5 |
0.711 |
|
48 |
36.3 |
35.5 |
39.7 |
36.5 |
1.9 |
0.498 |
|
72 |
47.5 |
46.5 |
44.7 |
43.0 |
2.0 |
0.719 |
|
Table 8. Relationship between RP level (kg DM/kg LW) and DM loss from grass incubated in the rumen of bulls for different times |
|||
|
Time, h |
Equation |
R2 |
P |
|
6 |
DM loss, % = 19.5 - 1.92 RP |
0.47 |
0.316 |
|
12 |
DM loss, % = 24.7 - 3.12 RP |
0.57 |
0.243 |
|
24 |
DM loss, % = 30.1 + 0.00 RP |
0.00 |
1.000 |
|
48 |
DM loss, % = 53.2 - 6.96 RP |
0.97 |
0.013 |
|
72 |
DM loss, % = 59.4 - 7.20 RP |
0.99 |
0.006 |
|
|
|
Figure 5. Relationship between RP level (kg DM/kg LW) and DM loss from grass incubated in the rumen of bulls for different times |
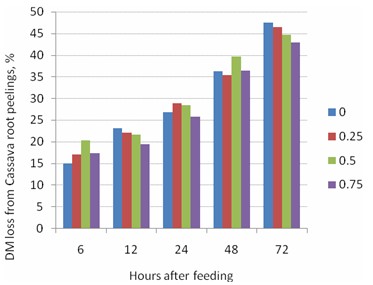
Effects of RP level on degradability of RP were less consistent (Table
7; Figure 6) and only significant for samples
taken after 72h when:
DM loss ,% = 47.7 – 6.12 RP; R2 = 0.99; P = 0.007
|
|
|
Figure 6. Relationship between RP level (kg DM/kg LW) and DM loss from cassava root peelings incubated in the rumen of bulls for different times |
Effects of the level of RP on OM degradability were similar to those for DM degradability (Table 9; Figure 7).
|
Table 9. Degradable organic matter (DOM) of feed samples after incubation |
||||||
|
After feeding, h |
RP0 |
RP0.25 |
RP0.5 |
RP0.75 |
SEM |
P |
|
Natural grass |
||||||
|
6 |
18.2 |
18.3 |
19.7 |
18.3 |
0.7 |
0.241 |
|
12 |
23.9 |
26.7 |
24.7 |
22.8 |
1.7 |
0.244 |
|
24 |
29.2 |
33.6 |
31.0 |
30.9 |
1.8 |
0.345 |
|
48 |
53.8 |
52.2 |
50.5 |
49.2 |
2.0 |
0.632 |
|
72 |
63.2 |
60.8 |
57.1 |
54.8 |
3.7 |
0.020 |
|
Cassava RP |
||||||
|
6 |
16.1 |
18.2 |
22.0 |
18.9 |
2.4 |
0.048 |
|
12 |
24.7 |
23.4 |
23.1 |
20.8 |
1.6 |
0.389 |
|
24 |
28.0 |
30.1 |
30.0 |
26.7 |
1.6 |
0.630 |
|
48 |
37.0 |
36.6 |
40.6 |
35.7 |
2.2 |
0.637 |
|
72 |
48.6 |
47.7 |
46.1 |
43.6 |
2.2 |
0.551 |
|
|
|
Figure 7. Relationship between diet RP level (kg DM/kg LW) and OM loss from grass incubated in the rumen of bulls for different times |
However, the trends were only significant for samples taken after 48 and 72h of incubation (Table 10).
|
Table 10. Relationship between diet RP level (kg DM/kg LW) and OM loss from grass incubated in the rumen of bulls for different times |
|||
|
Time, h |
Equation |
R2 |
P |
|
48 |
DM loss, % = 53.7 - 6.20 RP |
0.99 |
0.001 |
|
72 |
DM loss, % = 63.3 - 11.6 RP |
0.99 |
0.004 |
It is apparent that supplementing a basal diet of natural grass with dried cassava root peelings brought about changes in the rumen ecosystem.
The decrease in rumen bacteria and the increase in rumen protozoa populations in response to RP supplementation was unexpected. The increase in protozoa could reflect their affinity for the starch in the cassava RP, and by taking up a greater share of the rumen volume, it could be hypothesized that this would be at the expense of the rumen bacteria. The net result of such a change would be a less efficient rumen (Leng 1987). However, this interpretation is at odds with the finding that rumen ammonia levels were reduced, which implies enhanced growth of rumen microbes and therefore a greater supply of protein to the host animal. In turn, the observed increase in voluntary DM intake with RP supplementation is the expected response to an improved balance of nutrients that would result from an increase in rumen microbial growth (Preston and Leng 1987).
The possible effects of the cyanogenic glycosides on the rumen fermentation cannot be estimated in the absence of data on rumen levels of HCN. The estimated intakes of up to 2.1 mg HCN/kg live weight are at the lower end of the range of 2 to 4 mg/kg live weight considered to be toxic for cattle (Kumar 1992). However, Majak and Cheng (1984) fed cattle with Saskatoon serviceberry (Amelanchier alnifolia) (which contains the glycoside “runasin”) at levels equivalent to a HCN dose of 2.5 mg/kg live weight. They observed that the HCN produced in the rumen was rapidly metabolized by rumen microbes and there were no symptoms of toxicity in the animals. In an earlier experiment (Majak et al 1980) it was found that levels of the same plant needed to induce toxicity were equivalent to a HCN dose of 5.1 mg HCN equivalent/kg live weight. The fact that RP supplementgation led to increased feed intake indicates that there had been no deleterious effect on rumen fermentation or on the animal due to release of HCN in the rumen.
Supplementing a diet of natural grass fed to growing cattle with up to 0.75 kg DM/kg live weight of dried cassava root peelings:
Reduced the intake of the grass but increased by 10% the overall DM intake.
Reduced the concentration of rumen ammonia with minimal effects on rumen pH
Increased the rumen protozoal population but reduced that of bacteria
Reduced the rumen degradability of grass DM with minimal effects on the degradability of the cassava root peelings
It is concluded that there were unlikely to have been any negative effects on the rumen environment of the presence of cyanogenic glucosides in the cassava root peelings.
AOAC 1990 Official methods of analysis of the Association of Official Analytical Chemists (15th Edition.), Washington, DC. 1, 69-90.
Dehority B A 1984 Evaluation of subsampling and fixation procedures used for counting rumen protozoa. Applied Environment Microbiology 48 : 182-185
Devendra C 1997 Crop residues for feeding animal in Asia: Technology Development and Adoption in Crop/Livestock systems. In: Renard C (Editor) Crop Residues in Sustainable Mixed Crop/ Livestock Farming System, pp. 241–268
Doan Duc Vu 2004 Study on supplemented dried cassava leaf meal in the dairy ration based on rice straw. Reported Science of Nutrition and feed volume. Ha Noi 8-9/12/2004. Agricultural Publishing. Pp 179-187
Duong Nguyen Khang 2004 Cassava foliage as a protein source for cattle in Viet Nam. Doctoral thesis. Swedish University of Agricultural Sciences Uppsala
Duong Nguyen Khang and Wiktorsson H 2000 Effects of cassava leaf meal on the rumen environment of local yellow cattle fed urea-treated paddy straw. Asian –Australasian Journal of Animal Sciences 13: 1102-1108
Fernandez A and de B Hovell F D 1978 The rumen degradation of rice and cane fibre in animals eating sugar cane or molasses. Tropical Animal Production 3: 169 (abstract). http://www.fao.org/Ag/aga/AGAP/FRG/tap32/3_2_13.pdf
Gomez G and Valdivieso M 1988 The effects of ensiling cassava whole root chips on cyanide elimination. Nutrition Reports International 37: 1081-1092 and 1161-1166
Hill D C 1973 Chronic Cyanide Toxicity in Domestic Animals. p. 105-111. In Chronic cassava toxicity : Proceeding of an interdisciplinary workshop, London, England, 29-30 January 1973. International Development Research Centre Monography. IDRC-010e.
Kumar R 1992 Anti-nutritional factors, the potential risks of toxicity and methods to alleviate them. In Legume trees and other fodder tree as protein sources for livestock. A Speedy and P Pugliese (Editors). FAO, Animal Production and Health Paper. 102: 145-160 http://www.fao.org/Ag/AGA/AGAP/FRG/AHPP102/102-145.pdf
Leng R A 1987 Background nutrition digestive physiology, metabolism of ruminants. In: Tree foliage in ruminant nutrition. FAO Animal Production and Health Paper. 139: 8-27 http://www.fao.org/docrep/003/W7448E/W7448E00.htm
Majak W and Cheng K J 1984 Cyanogenesis in bovine rumen fluid and pure cultures of rumen bacteria. Journal of Animal Sciences 59: 784-790 http://jas.fass.org/cgi/reprint/59/3/784
Majak W, McDiarmid R E, Hall J W and Cheng K J 1990 Factors that determine rates of cyanogenesis in bovine fluid in vitro. Journal of Animal Sciences 68: 1648-1655 http://jas.fass.org/cgi/reprint/68/6/1648.pdf
Majak W, Udenberg T, Clark L J and McLean A 1980 Toxicity of Saskatoon Serviceberry to Cattle. Canadian Veterinary Journal 21 (3) 74-76
Man N V and Wiktorsson H 2001 Cassava tops ensiled with or without molasses as additive effects on quality, feed intake and digestibility by heifers. Asian-Australasian Journal of Animal Sciences 14 (5): 624-630
Minitab 2000 Minitab Reference Manual, PC Version, Release 12.2. Minitab Inc., State College, PA.
Nguyen Thi Thu Hong, Wanapat M, Wachirapakorn C, Padkee P and Rowlinson P 2003 Effects of timing of initial cutting and subsequent cutting on yields and chemical compositions of cassava hay and its supplementation on lactating dairy cows. Asian –Australasian Journal of Animal Sciences. 16: 1763-1769
Ørskov E R, de B Hovell F D and Mould F 1980 The use of the nylon bag technique for the evaluation of feedstuffs. Tropical Animal Production 5: (3): 195-213 http://www.fao.org/ag/AGA/AGAP/frg/tap53/53_1.pdf
Preston T R 1995 Tropical animal feeding- A manual for research worker. FAO. Animal production and health paper 126. Rome. http://www.fao.org/DOCREP/003/V9327E/V9327E00.HTM
Preston T A and Leng R A 1987 Matching Ruminant Production systems with Available Resources in the Tropics and Subtropics. Penambul Books, Armidales NSW http://www.utafoundation.org/P&L/preston&leng.htm
Ravindran V, Kornegay E T and Rajaguru A S B 1987 Influence of processing methods and storage time on cyanide potential of cassava leaf meal. Animal feed science and technology 17: 227-234
Ravindran V and Rajaguru A S B 1988 Effect of stem pruning on cassava root yield and leaf growth. Sri Lanka Journal of Agricultural Sciences 25: 32-37
Warner A C I 1962 Enumeration of rumen Micro- organisms. Journal of general Microbiology 28: 119-128
Received 9 July 2009; Accepted 4 August 2009; Published 1 September 2009