| Livestock Research for Rural Development 21 (8) 2009 | Guide for preparation of papers | LRRD News | Citation of this paper |
The effects of substituting hydrolysed feather meal (HFM) for freshwater shrimp meal (FSM; Caridina nilotica, Roux) on growth, digestibility and survival in Nile tilapia (Oreochromis niloticus L.) were evaluated under laboratory and practical culture conditions. Fingerlings with mean ranges of 26.0-26.5 and 35.4-37.0 g were held indoor in aquaria with recirculating water, and in cages that were installed in 800 m2 fertilized pond, respectively. Five isocaloric (12.3 kJ g-1) diets, containing between 28.5 and 31.0 % of crude protein, were prepared by gradually substituting HFM for FSM at rates of 0, 25, 50, 75 and 100% and fed to fish in aquaria; tilapia in cages were fed diets at substitution levels 0, 50 and 100%. All fish were fed at 10 % of body weight day-1 in three replicates for 84 days.
Results indicated that substitution of HFM for FSM at levels above 50% led to significant (P<0.05) growth reductions in aquaria. However, even 100% substitution rate did not significantly (P>0.05) affect growth of fish in the cages. In both experiments, survival was similar among treatments, but apparent protein digestibility decreased with increasing levels of HFM in the diet.
In conclusion, at least 50% substitution of HFM for FSM is possible in semi-intensive culture of Oreochromis niloticus, where natural food is available and where moderate growth performance levels may be accepted.
Key words: amino acids, aquaria, cage, nutrients, ponds
Production of farmed tilapia is among the fastest expanding food sectors in the world. Nile tilapia is the most cultured freshwater species among the farmed tilapias and contributes about 71 % of the total tilapia production (FAO 2002). However, the sustenance and expansion of production is limited by the high cost of fish feeds, which comprise over 50% of the production costs (Craig and Helfrich 2002). Protein usually is the most expensive nutrient and its level and quality determine the cost of fish feeds (Tacon 1993).
Most fish feeds contain a proportion of animal protein, which is considered superior to other sources in terms of palatability and nutrient availability (Tacon 1993; Hardy and Tacon 2002). The majority of fish feeds contain fish or shrimp meal as the main sources of dietary animal protein. However, due to worldwide decline in fishery products, fish and shrimp meals are increasingly becoming scarce and expensive (Mohsen and Lovell 1990). Consequently, nutritionists and feed manufacturers have been searching for alternative dietary animal protein from non-conventional sources to replace the conventional sources and reduce feed costs (El-Sayed 1998; Abdelghany 2003; Liti et al 2005). In Kenya, Omena (Rastrineobola argentea) and Freshwater Shrimp Meal (FSM) are commonly used as animal protein sources in fish feeds. However, Rastrineobola argentea is used for human food while FSM, a by-product of the Omena (Rastrineobola argentea) fishery in Lake Victoria is increasingly becoming scarce and expensive due to frequent closures of the lake’s fishery (Munguti et al 2006). Therefore, the replacement of FSM by cheaper and available animal protein feedstuffs is likely to contribute to reduce the costs of Oreochromis niloticus feeds.
Feather meal is high in protein (Tacon et al 1984), and could be incorporated into tilapia feeds after hydrolyzation. Previous studies have indicated that hydrolyzed feather meal (HFM) could only partially substitute fish meal without growth reduction in several fish species (Falaye 1982; Steffens 1994; Bishop et al 1995; Hasan et al 1997; Mendoza et al 2001). Improvement on the utilization of feather meal has mainly been based on two approaches, the direct supplementation of limiting amino acids (Murai et al 1982; Webster et al 1991), and the application of different techniques of hydrolyzation (Moran et al 1966; Tiews et al 1979; Hughes 1990; Steffens 1994; Mendoza et al 2001). However, direct amino acid supplementation may not be economically viable given their high cost and the low prices fetched by tilapias in developing sub-Saharan African countries. Furthermore, many investigators have expressed doubts on the viability of supplementing free amino acids in diets for tilapias, since free amino acids are assimilated long before the accompanying polypeptides, and before whole proteins are broken down; therefore, they are not presented at the same time for tissue formation (Jauncey 1982). On the contrary, tilapias are reported to utilize free dissolved amino acids in natural waters (Bowen 1980). Since Oreochromis niloticus feed continuously under semi-natural conditions (Getachew 1989), it is possible that a continuous supply of amino acids from the natural food web may have a potential in improving the nutritive value of feather meal under semi-intensive culture conditions.
The current study was conducted to analyse the effects of gradual substitution of freshwater shrimp meal by hydrolyzed feather meal on growth, apparent digestibility and survival of Oreochromis niloticus under semi-intensive culture conditions. Furthermore, potential differences in the response of fish kept in aquaria and those kept in cages under semi-intensive management should be recorded.
The study was conducted at Sagana Fish Farm (90 km northeast of Nairobi, altitude 1230 m, latitude 0°39´S and longitude 37°12´E). Two experiments were conducted, one in laboratory aquaria, each with a dimension of 0.45 x 0.3 x 0.3 m, and the other in net cages consisting of a top and bottom frames measuring 1.2 x 0.94 m and 0.9 x 0.9 m, respectively. Each cage had a height of 0.75 m and all cages were installed in a single earthen pond measuring 20 x 40 x 1.0 m. The aquaria were connected in a thermoregulated recirculating system, comprising a settling tank for solid removal and a tickling bio-filter to remove ammonia.
The pond was fertilized weekly at a rate of 20 kg N and 8 kg P ha-1 with urea and diammonium phosphate, respectively. Liming was done once at 2500 kg ha-1 with CaCO3 at the beginning of the experiment. Water quality was assessed by measuring temperature, pH, dissolved oxygen and chlorophyll a. The parameters were measured three times a week both in the aquaria and the cages. Dissolved oxygen was measured using model 57 oxygen meter (YSI industries, Yellow springs, OH, USA), while a glass electrode-pH meter, Hi-9024 microcomputer (Hanna Instruments Ltd., Chicago, IL, USA), was used to measure pH. Chlorophyll a was determined as described in APHA (1990).
Fresh chicken feathers were collected from hotels around Sagana town and were sun-dried. The feathers were ground using a hammer mill and were then mixed with a solution of NaOH (5 g/l) at a ratio of 0.5 l per 1 kg of feathers. Thereafter, ground feathers were hydrolysed in an autoclave at 16 psi and about 110°C for 3 hours. The hydrolysed feather meal was sun-dried and further ground into a fine powder using a blender. A batch of freshwater shrimp was purchased from supply stores in Kisumu, and cotton seed cake from a local feed manufacturer. Wheat bran was purchased from a local wheat-processing factory. The freshwater shrimp and wheat bran were ground using a hammer mill.
The processed ingredients were subjected to proximate analysis before formulation of experimental diets. The analysis was carried out as described in AOAC (1990) and in triplicates. Feedstuffs were analysed for crude protein (CP), ether extracts (EE), ash, nitrogen-free extracts (NfE), and crude fibre (CF). CP was estimated from Kjeldahl nitrogen by multiplication with a factor of 6.25, while EE were quantified through the loss in weight after extraction of the sample with petroleum ether (boiling point 40-60oC). Ash was determined by ashing dry samples in a muffle furnace at 550oC for 4 hours. CF was determined as the ash-free residues of a digestion process in alkaline and acidic solution. NfE was determined by the difference method (dry matter-CP-EE-CF-Ash). The proximate composition of feed ingredients is shown in Table 1.
|
Table 1. Proximate composition of the feedstuffs used in diet formulation (as-fed basis) |
||||||
|
Ingredienta) |
Nutrientb), % |
|||||
|
DM |
CP |
EE |
CF |
NfE |
Ash |
|
|
FSM |
87.5 |
60.3 |
1.4 |
6.2 |
6.7 |
24.8 |
|
HFM |
89.9 |
80.7 |
1.8 |
3.2 |
4.2 |
10.1 |
|
CSC |
89.8 |
34.9 |
12.8 |
25.8 |
19.4 |
6.0 |
|
WB |
88.0 |
14.0 |
5.9 |
13.6 |
60.2 |
6.3 |
|
a)FSM = freshwater shrimp meal, HFM = hydrolysed feather meal; CSC = cotton seed cake; WB = wheat bran b)DM = dry matter, CP = crude protein, EE = ether extracts, CF = crude fibre, NfE = nitrogen-free extracts |
||||||
Five experimental diets, each formulated to contain 30 % of CP and 12.3 kJ g-1 digestible energy were produced by including 0, 2.25, 4.5, 6.75 and 9.4 % of HFM, together with small amounts of Carboxyl methyl cellulose as a filler material instead of FSM, thereby representing substitution rates of 0, 25, 50, 75 and 100%, respectively (Table 2).
|
Table 2. Composition and results of proximate analysis (as-fed basis) of experimental diets |
|||||
|
Ingredients / Proximate analysis |
Dietary treatment (% of fresh-water shrimp meal substituted by hydrolysed feather meal) |
||||
|
0% (control) |
25% |
50% |
75% |
100% |
|
|
Freshwater shrimp meal, % |
12 |
9 |
6 |
3 |
0 |
|
Hydrolysed feather meal, % |
0 |
2.25 |
4.5 |
6.75 |
9.4 |
|
Cotton seed cake, % |
25 |
25 |
25 |
25 |
25 |
|
Wheat bran, % |
63 |
63 |
63 |
63 |
63 |
|
Carboxyl methyl cellulose, % |
0 |
0.75 |
1.5 |
2.25 |
2.6 |
|
Dry matter, % |
88.5 |
88.7 |
90.1 |
89.2 |
90.3 |
|
Crude protein, % |
28.5 |
29.1 |
29.8 |
30.4 |
31.0 |
|
Ether extracts, % |
5.6 |
6.6 |
4.6 |
5.6 |
6.6 |
|
Crude fibre, % |
14.8 |
16.7 |
16.6 |
15.5 |
14.4 |
|
NfE, % |
43.2 |
40.1 |
42.0 |
41.9 |
41.8 |
|
Ash, % |
7.9 |
7.5 |
7.0 |
6.6 |
6.2 |
|
DE, kJ g-1 |
12.3 |
12.3 |
12.3 |
12.3 |
12.3 |
|
DE = digestible energy; NfE = Nitrogen Free Extracts |
|||||
Each ingredient was homogeneously ground to pass through a 200mm sieve. The formulations were made by mixing the necessary ingredients into a homogenate, which was moistened before passing through a modified meat mincer. The resulting expeller-like strands were sun-dried and stored at room temperature. The amino acid profiles of feed ingredients are shown in Table 3.
|
Table 3. Amino acid content of feed ingredients (% on as-fed basis) |
||||||||||||
|
Feed ingredient |
His |
Thr |
Arg |
Tyr |
Phe |
Val |
Ile |
Leu |
Lys |
Cys |
Met |
Trp |
|
FSM |
1.1 |
2.3 |
4.3 |
1.9 |
2.3 |
2.7 |
2.4 |
3.9 |
3.4 |
0.6 |
1.4 |
0.6 |
|
HFM |
0.4 |
5.1 |
7.7 |
2.4 |
4.8 |
7.7 |
4.9 |
8.3 |
1.3 |
3.2 |
0.3 |
0.4 |
|
CSC |
0.9 |
1.2 |
1.1 |
1.0 |
1.6 |
1.5 |
1.1 |
2.1 |
1.6 |
0.6 |
0.6 |
0.4 |
|
WB |
0.4 |
0.6 |
1.1 |
0.5 |
0.7 |
0.8 |
0.6 |
1.1 |
0.5 |
0.4 |
0.3 |
0.2 |
|
FSM = freshwater shrimp meal, HFM = hydrolysed feather meal; CSC = cotton seed cake; WB = wheat bran |
||||||||||||
The diets were randomly allocated to groups of hand-sexed male Oreochromis niloticus fingerlings that were held in aquaria and cages. Those in aquaria were fed all the five diets in five treatments, while the caged fish were fed diets at substitution levels of 0, 50 and 100% in three treatments; both experiments had three replicates per treatment. The initial average stocking weight in aquaria and cages were 26 and 36g, respectively, at densities of 6 and 15 fish, respectively. Fish were acclimatised for two weeks prior to the start of each experiment. Feed was offered at 10% of body weight per day for 84 days. Sampling was done on a bi-weekly basis to monitor growth and adjust feeding rates. Fish were fed using the automated feeders described by Waidbacher et al (2006). The feeders were calibrated to deliver feed continuously between 8.00 and 18.00 hrs. At the end of the study, all fish from cages and aquaria were harvested, weighed and counted.
Final harvest mean weight, weight gain, specific growth rate and apparent protein digestibility coefficient were measured. Specific growth rate (SGR) was calculated using the following equation:
Where Wi and Wf are the initial and final mean body weights, respectively and t is time in days from stocking to harvest.
The apparent digestibility coefficient for protein (ADCp) was determined using acid-insoluble ash (AIA) as the indigenous inert marker. The following formula was applied:
![]() (adjusted
from Maynard and Loosli 1962)
(adjusted
from Maynard and Loosli 1962)
Where CPf is faecal CP, CPd is the dietary CP; AIAd is the marker in the diet and AIAf is the marker in the faeces. The ash content was analyzed by ashing the sample in a furnace at 550°C for 6 hours. The residue was then treated with HCl (2.5 %) and afterwards was ashed again for 2 hours, the residue representing acid-insoluble ash (AOAC 1990).
Statistical analysis was performed using SPSS (2006) statistical software package. Data were subjected to a one-way analysis of variance. Where appropriate, data were transformed to conform to the requirements for the test. Duncan multiple range test was applied to identify means that were significantly different from each other (Duncan 1955). The relationship between HFM inclusion level and ADCp was evaluated by applying linear regression analysis as described by Sokal and Rohlf (1981). A type I error of 0.05 was used to define significance.
Water quality parameters did not vary significantly (P≥0.05) among treatments, both in cages and aquaria during the experimental period. Mean value for chlorophyll a was 162.4±17.1 mg m-3. The mean ranges for dissolved oxygen values in the cages were 3-10 mg L-1. (mean: 6.2 mg L-1.). Water temperature in cages ranged from 24.3 to 28.9 oC (mean: 27.0±0.5 oC) in the pond,while the water temperature was maintained between 26 and 28oC (mean: 27.0±0.04oC) in the aquaria. The pH values ranged from 7.8 to 8.3 (mean: 8.1±0.03) in the experimental pond and 7.0-7.5 (mean: 7.2±0.01) in the aquaria. In all cases, water quality parameters were within the values recommended for tilapia culture.
Data on fish growth performance for aquaria and cages are presented in Table 4 and 5, respectively.
|
Table 4. Growth performance of Oreochromis niloticus fed diets containing increasing levels of HFM in substitution for FSM in aquaria |
||||||
|
Parameter |
Dietary treatment (rate of substitution for FSM) |
SE |
||||
|
0 % |
25 % |
50 % |
75 % |
100 % |
||
|
Initial body weight, g |
26.5a |
26.0a |
26.2a |
26.5a |
26.2a |
0.35 |
|
Final body weight, g |
43.4a |
41.9a |
41.2a |
34.2b |
33.4 b |
0.91 |
|
Weight gain, % |
67.1a |
61.3a |
58.3a |
31.4b |
28.4b |
3.51 |
|
Specific growth rate, % |
0.61a |
0.57a |
0.54a |
0.32b |
0.30b |
0.03 |
|
Survival, % |
97.6a |
95.3 a |
95.3a |
95.3a |
100a |
1.91 |
|
ab means in the same row for each parameter with different superscripts are significantly different (p <0.05) SE = standard error of mean |
||||||
|
Table 5. Growth performance of Oreochromis niloticus fed diets containing increasing levels of HFM in substitution for FSM in cages |
||||
|
Parameter |
Dietary treatment (rate of substitution for FSM) |
SE |
||
|
0 % |
50 % |
100 % |
||
|
Initial body weight, g |
37.0a |
35.4a |
36.0a |
1.00 |
|
Final body weight, g |
83.2a |
82.2a |
77.5a |
3.40 |
|
Weight gain, % |
131.2a |
128.2a |
115.7a |
9.44 |
|
Specific growth rate, % |
0.99a |
0.98a |
0.91a |
0.05 |
|
Survival, % |
97.6a |
95.3a |
100a |
1.91 |
|
a means in the same row for each parameter with identical superscript are not significantly different (p ≥ 0.05) SE = standard error of mean |
||||
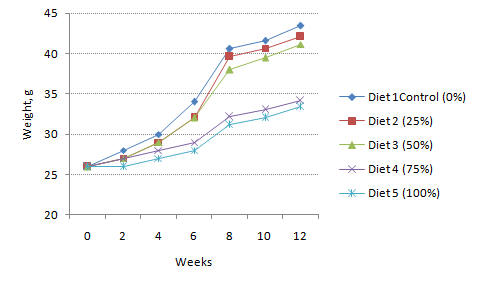
Fish that were fed diets containing 0, 25 and 50% HFM in aquaria had similar mean weight, specific growth rate and percent weight gain and grew significantly better (P<0.05) than those fed diets in which 75 and 100% of FSM were substituted for HFM. However, there were no significant differences (P≥0.05) in growth performance between the 0, 50 and 100% HFM diet in the cages. Survival rate was high and very similar (P>0.05) among treatments both in aquaria and cages. Growth curves for Oreochromis niloticus in aquaria and cages are presented in Figures 1 and 2, respectively.
|
|
|
Figure 1. Growth curves for Oreochromis niloticus receiving diets containing different levels of HFM in aquaria |
|
|
|
Figure 2.
Growth curves for Oreochromis niloticus receiving diets |
After about two weeks from the time of stocking, fish receiving diets in which 0 to 50% of FSM had been substituted for HFM differed from the other treatments in the aquaria experiment.
Data on apparent protein digestibility (ADCp) of fish in aquaria and cages are shown in Table 6 and Figure 3.
|
Table 6. Apparent protein digestibility coefficient (APDCp, %) in cages and aquaria |
||||||
|
Parameter |
Dietary treatments |
SE |
||||
|
0% |
25% |
50% |
75% |
100% |
||
|
ADCp( cage) |
64.5a |
- |
64.0a |
- |
54.6b |
0.39 |
|
ADCp (aquaria) |
62.2a |
58.0b |
52.2c |
46.5d |
34.7e |
0.96 |
|
abcde means in the same row for each parameter with different superscripts are significantly different (p <0.05) - 25 and 75 % diets were fed in aquaria only SE = standard error of mean |
||||||
|
|
|
Figure 3. Apparent
protein digestibility (observed vs. estimated) |
There were significant differences (P<0.05) in ADCp between all treatments in the aquaria experiment. The ADCp decreased with increasing levels of HFM both in aquaria and cages, with regression slopes of -0.26±0.02 and -0.098±0.02, respectively. In cages, only the 100 % treatment showed reduced ADCp.
Data from the present study indicate that the growth of groups of Oreochromis niloticus that were fed diets containing a mixture of freshwater shrimp (FSM) and hydrolyzed feather meal (HFM) protein in cages was similar to those fed FSM as the only source of animal protein (control treatment). However, growth significantly declined when fish were fed diets containing over 4.5 % of HFM (i.e. 75-100 % FSM-substitution level) in aquaria.
The differences in growth performance between the caged and aquaria fish in the present study may be attributed to different culture conditions. The aquaria experiment in the present study took place in indoor laboratory environments, while the cage experiment was conducted in a fertilized earthen pond. This setup probably allowed for some extra nutrition from natural food in the pond, which may have supplemented potentially deficient dietary amino acids. HFM has been reported to be deficient in certain essential amino acids (Falaye 1982; Tacon et al 1984; Mendoza et al 2001); therefore, the interaction between the components of pond’s natural food and feather meal may have improved the essential amino acid supply from the latter, thus resulting in better utilization and growth performance of fish in the cages. It should however be noted that these results were derived at an overall moderate level of performance, which can be considered as typical for the given setting, representing semi-intensive production under East African conditions.
The findings from the aquaria experiment are similar to those reported by other investigators. Falaye (1982) did not observe a significant decrease in growth parameters of Oreochromis niloticus when 50 % of fish meal was substituted for HFM. Similar results were reported by Viola and Zohar (1984), who concluded that up to 50 % of fish meal could be replaced by poultry by-products in diets of tilapia hybrids without deleterious effects on fish growth. However, Bishop et al (1995) reported a slightly higher substitution level (66 %) of fish meal protein for HFM protein in diets for Oreochromis niloticus without significant reduction in growth. The higher substitution level reported by these authors as compared to that from the aquaria experiment of the present study may be attributed to differences in culture conditions. The present experiment was conducted in aquaria, which were cleaned every day, while that of Bishop et al (1995) was conducted in concrete tanks where some development of autotrophs and heterotrophs was possible.
The decrease in growth observed in fish fed diets containing over 4.5 % HFM (i.e. substitution rates above 50 %) and kept in aquaria may be due to sub-optimal intake of essential amino acids (Jauncey and Ross 1982; NRC 1983; Santiago and Lovell 1988) from HFM, which might have hindered proper utilization of dietary HFM protein and its subsequent synthesis to body protein. Attempts have been made to improve the nutritive value of feather meal-based diets for some aquatic species by supplementation of deficient amino acids (Tiews et al 1979; Hughes 1990; Steffens 1994). However, Falaye (1982) reported a decreased performance of fish growth when HFM contributed over 50% of the animal protein even after some deficient amino acids were supplemented. The author suggested that growth inhibition might have involved factors other than amino-acid imbalances.
Reports on utilization of supplementary amino acids are variable and conflicting, with some authors reporting improved growth of fish after supplementation (Murai et al 1982; Webster et al 1991), while others report no response in growth (Ng and Wee 1979; El-Sayed 1990). Other investigators have suggested that the utilization of amino acids is dependent on meal frequencies (Webster et al 1991): channel catfish fed once a day, may not sufficiently utilize supplemented synthetic amino acids for protein synthesis, but growth was improved to the level of the control when diets containing L-lysine were fed twice a day. Under natural or semi-natural conditions, Oreochromis niloticus feed continuously during day time (Getachew and Fernando 1989; Zenebe and Getachew 1998), thus based on the hypothesis of meal frequency, utilization of free amino acids might be enhanced. Bowen (1980) reported that tilapias can thrive well on naturally occurring dissolved amino acids. This mode of feeding would ensure that free amino acids were supplied continuously at the site of tissue formation. The observation by Bowen (1980) and the lack of differences between the control and the diets containing HFM in the cages in growth performance in the present study suggest that the supply of amino acids from natural food may be an economically attractive strategy of supplementing limiting amino acids in tilapia diets.
The method used to process feathers in the present study produces two categories of essential amino acids, a batch of free amino acids and those contained in the remaining polypeptides and intact protein. The set of free essential amino acids is likely to be irreversibly absorbed before those in the polypeptides and intact protein are liberated for tissue protein synthesis (Yamada et al 1981; Cowey and Walton 1988), eventually leading to poor utilization of feather meal. However, the continuous supply of amino acids from natural food could enhance utilization of feather meal by upgrading the profile of both free essential amino acids, and those contained in polypeptides and protein after liberation.
In the present study, ADCp declined with increasing levels of HFM in the diets. This trend agrees with that reported by Degani et al (1997) and Yang et al (2006), who observed a similar decrease in ADCp with increasing levels of feather meal in Carp (Cyprinus carpio L.) and gibel carp (Carassius auratus gibelio Bloch), respectively. The regression slope for apparent protein digestibility (b=-0.098) of feather meal based diets in the cage experiment was less negative than the values calculated for fish in aquaria (b=-0.26), suggesting a less pronounced depression of protein digestibility in a pond environment. Several factors emerging from the pond environment may have contributed to these differences: Algae or algal based diets are reported to stimulate secretion of copious amounts of gastric acid in Oreochromis niloticus and Oreochromis mossambicus (Bowen 1981; Getachew 1987, 1989); this may have led to a more acidic medium and eventually in an enhanced ADCp. On the other hand, ADCp in the cage environment was probably even systematically underestimated herein, as it was technically not feasible to properly account for the additional intake of natural food in the digestibility study.
In conclusion, the present study demonstrated that hydrolyzed feather meal could substitute freshwater shrimp meal at least up to 50% in the diets of Oreochromis niloticus if the level of growth performance aimed at is below maxiumum. If fish have the chance to feed on materials from a pond under semi-intensive culture conditions, an even higher substitution rate may be possible without compromising growth.
The authors thank the Austrian Development Cooperation through ÖAD North-South Dialogue scholarship programme and the Department of Water, Atmosphere and Environment of BOKU-University of Natural Resources and Applied Life Sciences Vienna, for providing financial support to this project. Members of Sagana Aquaculture Center and Kenya Marine and Fisheries Research Institute (KMFRI) are acknowledged for their immense support to the project. We are grateful to Silke Drexler for her help in the preparation of this paper.
Abdelghany A E 2003 Partial and complete replacement of fishmeal with gambusia meal in diets of red tilapia (Oreochromis niloticus x O.mossambicus). Aquaculture Nutrition 9:145-151
AOAC 1990 Official Methods of Analysis of the Association of Official Analytical Chemists, Arlington, VA, USA, 1018p.
APHA 1990 Standard Method for Examination of Water and Wastewater, 17th edition. pp. 10-203. American Public Health Association, Washington, DC
Bishop C D Angus R A and Watts A 1995 The use of feather meal as a replacement for fishmeal in the diet of Oreochromis niloticus fry. Bioresource Technology 54: 291-295
Bowen S H 1980 Detrital Non-protein Amino Acids are the Key to Rapid Growth of Tilapia in Lake Valencia, Venezuela. Science 207: 1216-1218
Bowen S H 1981 Digestion and assimilation of periphytic detrital aggregrate by Tilapia mossambica. Transactions of the American Fisheries Society 110: 239-245
Cowey C B and Walton M J 1988 Studies on the uptake of (14C) amino acids derived from both dietary (14C) protein and dietary (14C) amino acids by rainbow trout, Salmo gairdneri Richardson. Journal of Fish Biology 33: 293-305
Craig S and L A Helfrich 2002 Understanding Fish Nutrition, Feeds and Feeding. Cooperative Extension Service publication 420-256. Virginia State University, USA http://pubs.ext.vt.edu/420/420-256/420-256.pdf
Degani G, Viola S and Yehuda Y 1997 Apparent digestibility coefficient of protein sources for carp, Cyprinus carpio L. Aquaculture Research 28: 23-28
Duncan D B 1955 Multiple Range and Multiple F-Tests. Biometrics 11: 1-42
El-Sayed A F M 1990 Long-term evaluation of cotton seed meal as a protein source for Nile tilapia, Oreochromis niloticus. Aquaculture 84: 315-320
El-Sayed A F M 1998 Total replacement of fishmeal with animal protein sources in Nile tilapia Oreochromis niloticus (L) feeds. Aquaculture Research 29: 275-280.
Falaye A E 1982 The use of hydrolysed feather meal alone or in combination with supplemental amino acids as a dietary source for tilapia (Oreochromis niloticus). MSc thesis, University of Stirling, UK
FAO 2002 Global Aquaculture Production. Fishery Statistical Collections. FAO Fisheries and Aquaculture Department, Rome, Italy. http://www.fao.org/fishery/culturedspecies/Oreochromis_niloticus#tcN90112
Getachew T 1987 A study on an herbivorous fish (Oreochromis niloticus L.) diet and its quality in two Ethiopian Rift valley lakes, Awasa and Zwai. Journal of Fish Biology 30: 439-449
Getachew T 1989 Stomach pH, feeding rhythm and ingestion rate in Oreochromis niloticus L. (Pisces: Cichlidae) in Lake Awasa, Ethiopia. Hydrobiologia 174: 43-48
Getachew T and Fernando C H 1989 The food habits of a herbivorous fish (Oreochromis niloticus L.) in Lake Awasa, Ethiopia. Hydrobiologia 194: 195-200
Hardy R W and Tacon A D J 2002 Fish meal: historical uses, production trends and future outlook supplies. In: Stickney R R and MacVey J P (editors), Responsible Marine Aquaculture, CABI Publishing, New York, USA, pp. 311-325
Hasan M R, Haq M S, Das P M and Mowlah G 1997 Evaluation of poultry-feather meal as a dietary protein source for Indian major carp, Labeo rohita fry. Aquaculture 151: 47-54
Hughes S 1990 Feather meal can displace fishmeal in aquaculture rations. Feed international 15: 13-15
Jauncey K 1982 The effect of varying dietary protein level on growth, food conversion and protein utilization and body composition of juvenile tilapia (Sarotherodon mossambicus). Aquaculture 27: 43-54
Jauncey K and Ross B 1982 A guide to tilapia feeds and feeding. Institute of Aquaculture, University of Stirling, Scotland. U.K, 111 p.
Liti D, Kerogo L, Munguti J and Chhorn L 2005 Growth and economic performance of Nile tilapia (Oreochromis niloticus L.) fed on two formulated diets and two locally available feeds in fertilized ponds. Aquaculture Research 336: 746-752
Maynard A L and Loosli K J 1962 Animal nutrition, 5th edition, McGraw-Hill, New York, 613 p.
Mendoza R, De Dois A, Vazquez C, Cruz E, Ricque D Aguilera C and Montemayor J 2001 Fishmeal replacement with feather-enzymatic hydrolyzates co-extruded with soya-bean meal in practical diets for pacific white shrimp (Litopenaeus vannamei). Aquaculture Nutrition 7: 143-151.
Mohsen A A and Lovell E T 1990 Partial substitution of soybean meal with animal protein sources in diets for channel catfish. Aquaculture 90: 303-311
Moran E, Summers J and Slinger S 1966 Keratin as a source of protein for the growing chick. Poultry Science 45: 1255-1257
Munguti J M, Liti D M, Waidbacher H, Straif M and Zollitsch W 2006 Proximate composition of selected potential feedstuffs for Nile tilapia (Oreochromis niloticus Linnaeus) Production in Kenya. Die Bodenkultur – Austrian Journal of Agricultural Research 57: 123-133
Murai T, Ogata H and Nose T 1982 Methionine coated with various minerals supplemented to soybean meal diet for fingerling carp, Cyprinous carpio and channel catfish Ictarulus punctatus. Bulletin of the Japanese Society of Scientific Fisheries 48: 85-88
NRC (National Research Council) 1983 Nutrient requirement of warm water fishes and shellfishes, National Academic Press, Washington, D.C., USA
Ng W K and Wee K L 1979 The nutritive value of cassava leaf meal in pelleted feed for Nile tilapia. Aquaculture 83: 45-58
Santiago C B and Lovell R T 1988 Amino acid requirement for growth of Nile tilapia. The Journal of Nutrition 118: 1540-1546 http://jn.nutrition.org/cgi/reprint/118/12/1540.pdf
Sokal R R and Rohlf F J 1981 Biometry, The principles and practice of statistics in biological research, W H Freeman and Company, San Francisco, USA
SPSS 2006 SPSS for Windows 14.0, SPSS Inc., Chicago, Illinois, USA
Steffens W 1994 Replacing fishmeal with poultry by product meal in diets of rainbow trout (Onchorhynchus mkiss). Aquaculture 124: 27-34
Tacon A G J 1993 Feed ingredients for warm water fish, Fishmeal and other processed feedstuffs. FAO Fisheries Circular No.856, FAO Rome, 64 p.
Tacon A G J, Jauncey K, Falaye A, Pantha M, McGowen I. and Stafford E 1984 The use of meat and bone meal, hydrolyzed feather meal and soyabean meal in practical fry and fingerling diets for Oreochromis niloticus. In: Fishelson L and Yaron Z (editors), Proceedings of the First International Symposium on Tilapia in Aquaculture, Tel Aviv University Press, Tel Aviv, Israel, pp. 356-365
Tiews K, Koops H, Gropp J and Beck H 1979 Compilation of fishmeal free diets obtained in rainbow trout (Salmo gairdneri) feeding experiments at humburg (1970-77/78). In: Halver J E and Tiews K (editors), Finfish Nutrition and Fish Feed Technology, Hamburg, Berlin, Germany, pp. 219-228
Viola S and Zohar G 1984 Nutrition studies with market size hybrids of tilapia (Oreochromis niloticus) in intensive culture. The Israeli Journal of Aquaculture-Bamidgeh 36: 3-15
Waidbacher H, Liti D M, Fungomeli M, Mbaluka R K, Munguti J M and Straif M 2006 Influence of pond fertilization and feeding rate on growth performance, economic returns and water quality in a small-scale cage cum-pond integrated system for production of Nile tilapia (Oreochromis niloticus L.). Aquaculture Research 37: 594-600
Webster C D Tidwell J H and Yancey D H 1991 Evaluation of distillers’ grains with solubles as a protein source in diets for channel catfish. Aquaculture 96: 179-190
Yamada S, Simpson K, Tanaka Y and Katayama T 1981 Plasma amino acid changes in rainbow trout force-fed casein and corresponding amino acid mixture. Bulletin of the Japanese Society of Scientific Fisheries 47: 1035-1040
Yang Y, Xie S, Cui Y, Zhu X, Lei W and Yang Y 2006 Partial and total replacement of fishmeal with poultry by-products meal in diets of gibel carp, Carassius auratus gibelio Blouch. Aquaculture Research 37: 40-48
Zenebe T and Getachew T 1998 Diel feeding rhythm and assimilation efficiency of Oreochromis niloticus L, Pisces: Cichlidae) in Lake Ziway, Ethiopia. Verhandlungen - Internationale Vereinigung für theoretische und angewandte Limnologie 26: 2324-2328
Received 24 February 2009; Accepted 8 May 2009; Published 5 August 2009